Hydrodynamic and Soil Biodiversity Characterization in an Active Landslide

 ,
,  ,
,  , ,
, ,
Abstract
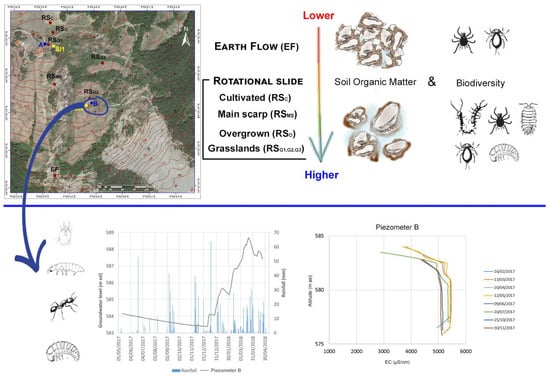
:
1. Introduction
2. Materials and Methods
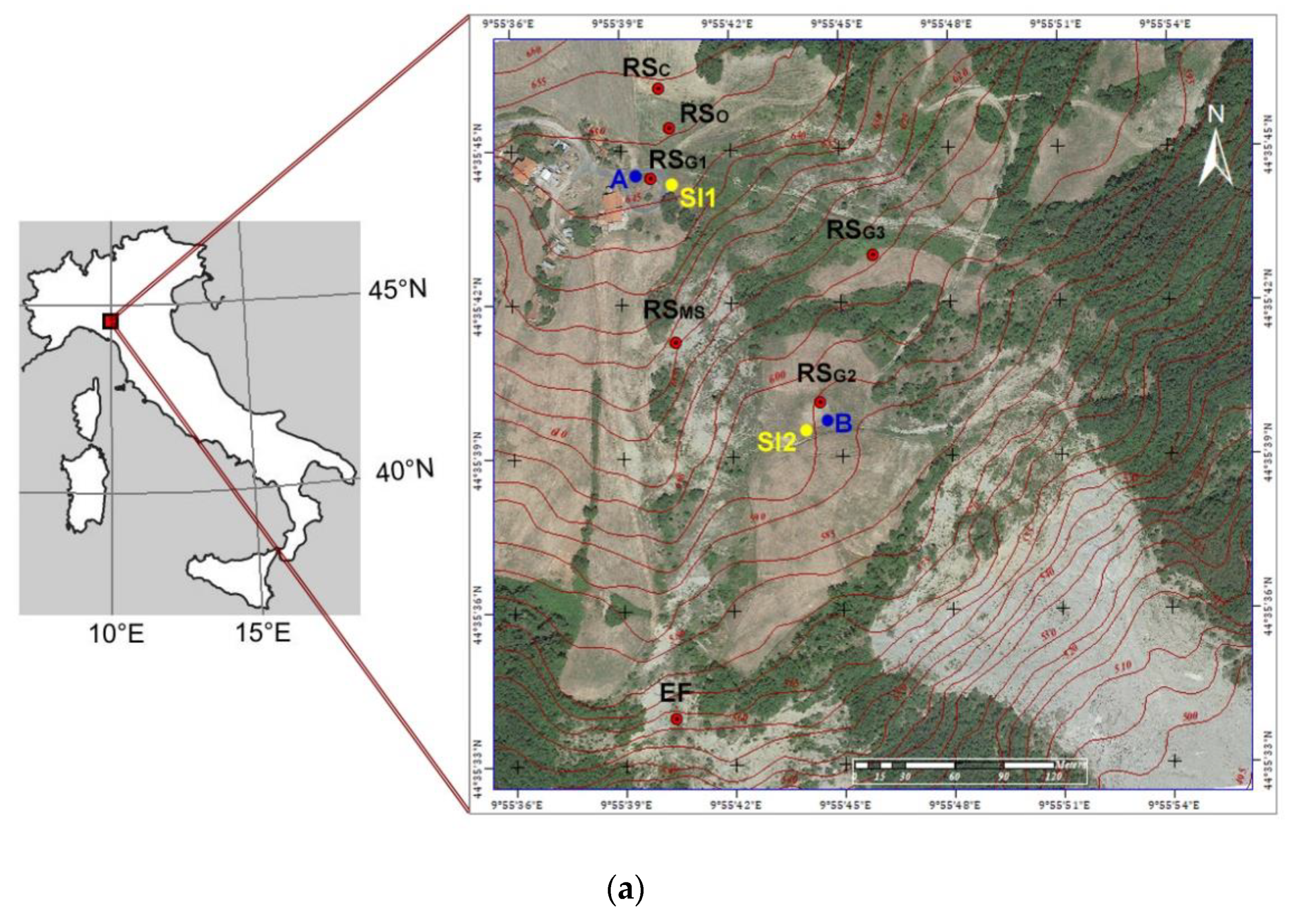
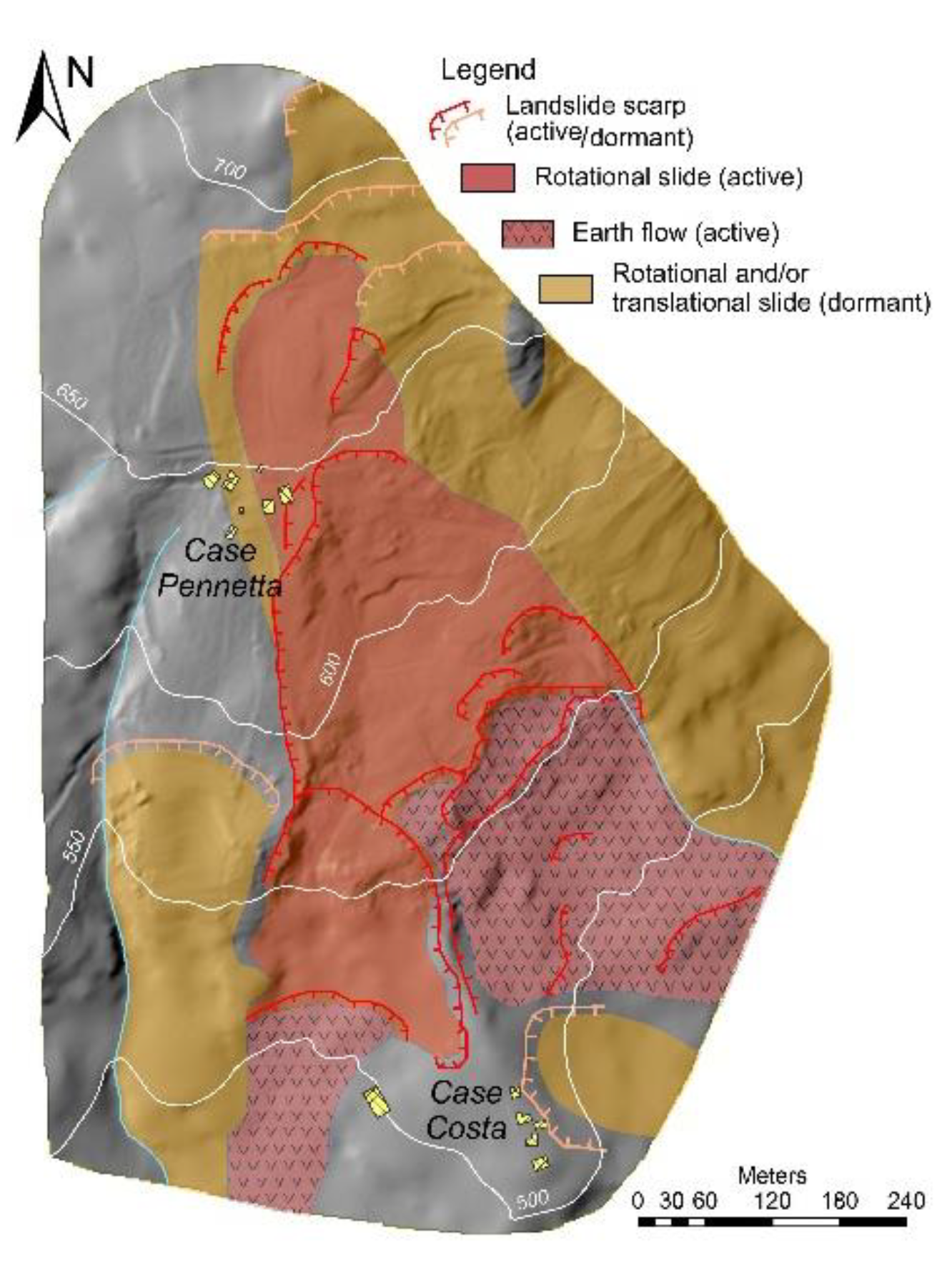
2.1. Study Area
2.2. Geological and Geomorphological Survey
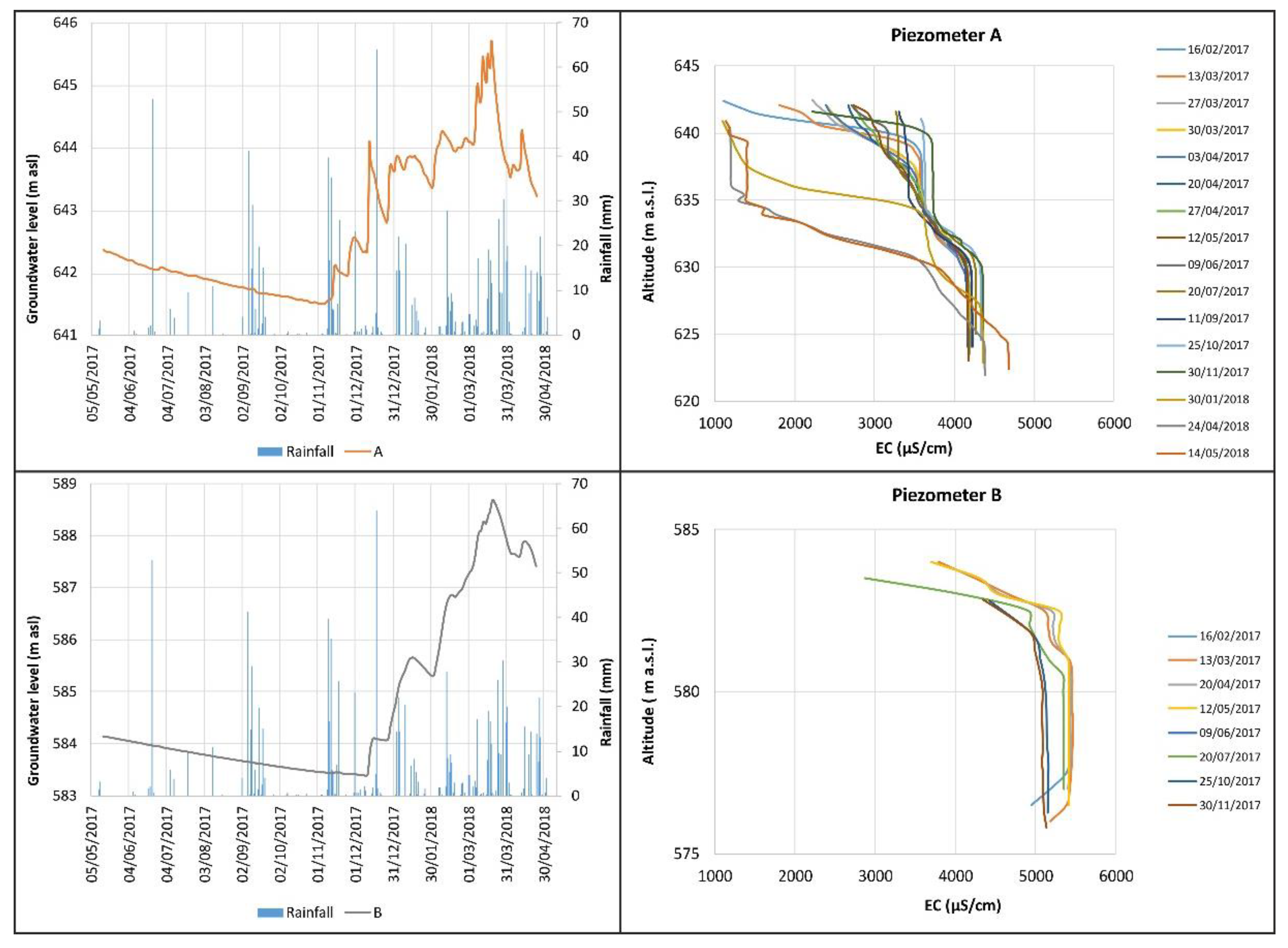
2.3. Hydrogeological Investigations
2.4. Soil Samples and Chemical Analysis
2.5. Soil Arthropod Investigation
2.6. Statistical Analysis
3. Results
3.1. Geological and Geomorphological Features of the Landslide
3.2. Hydrogeological Behaviour
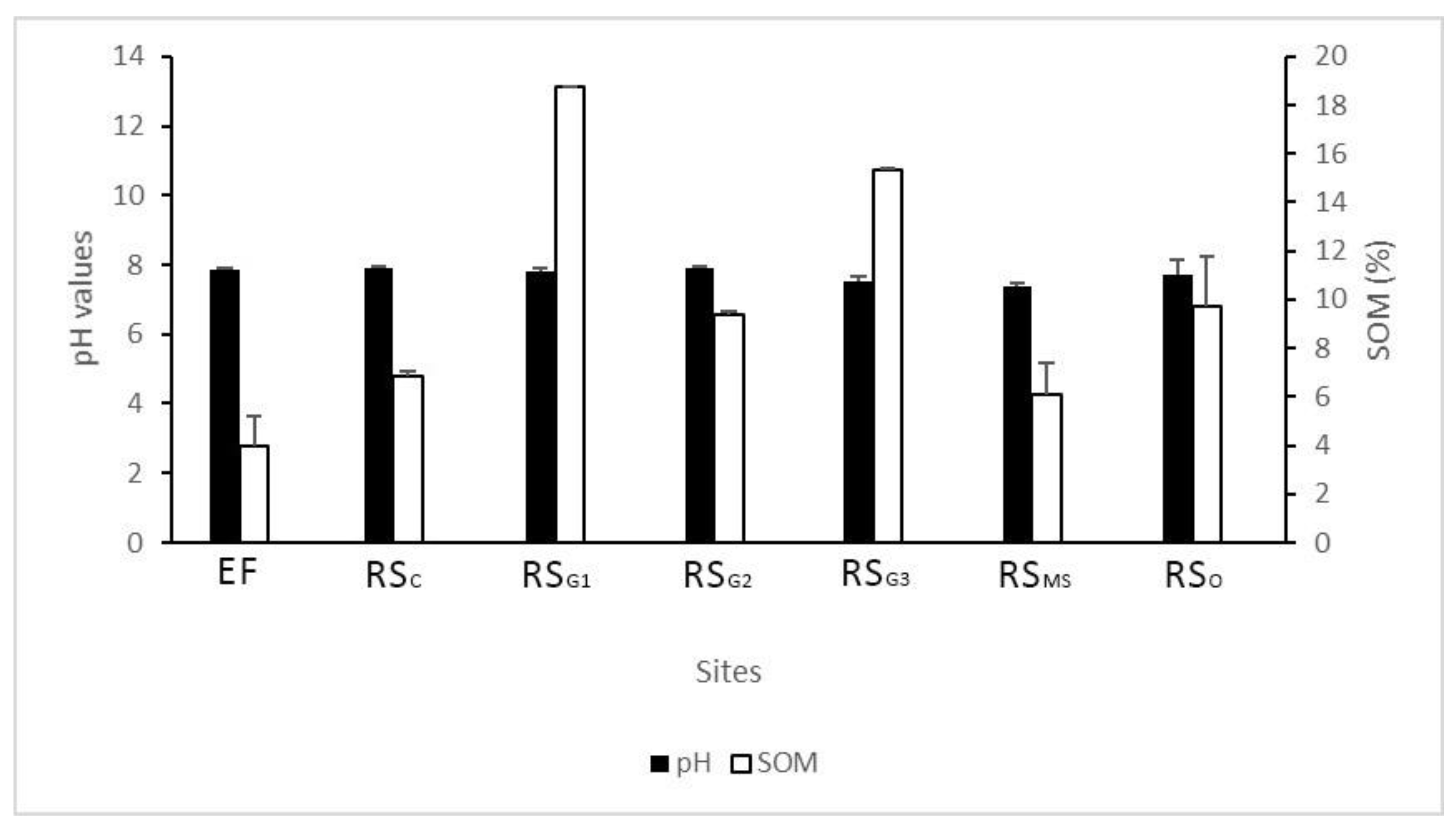
3.3. Soil Characterization
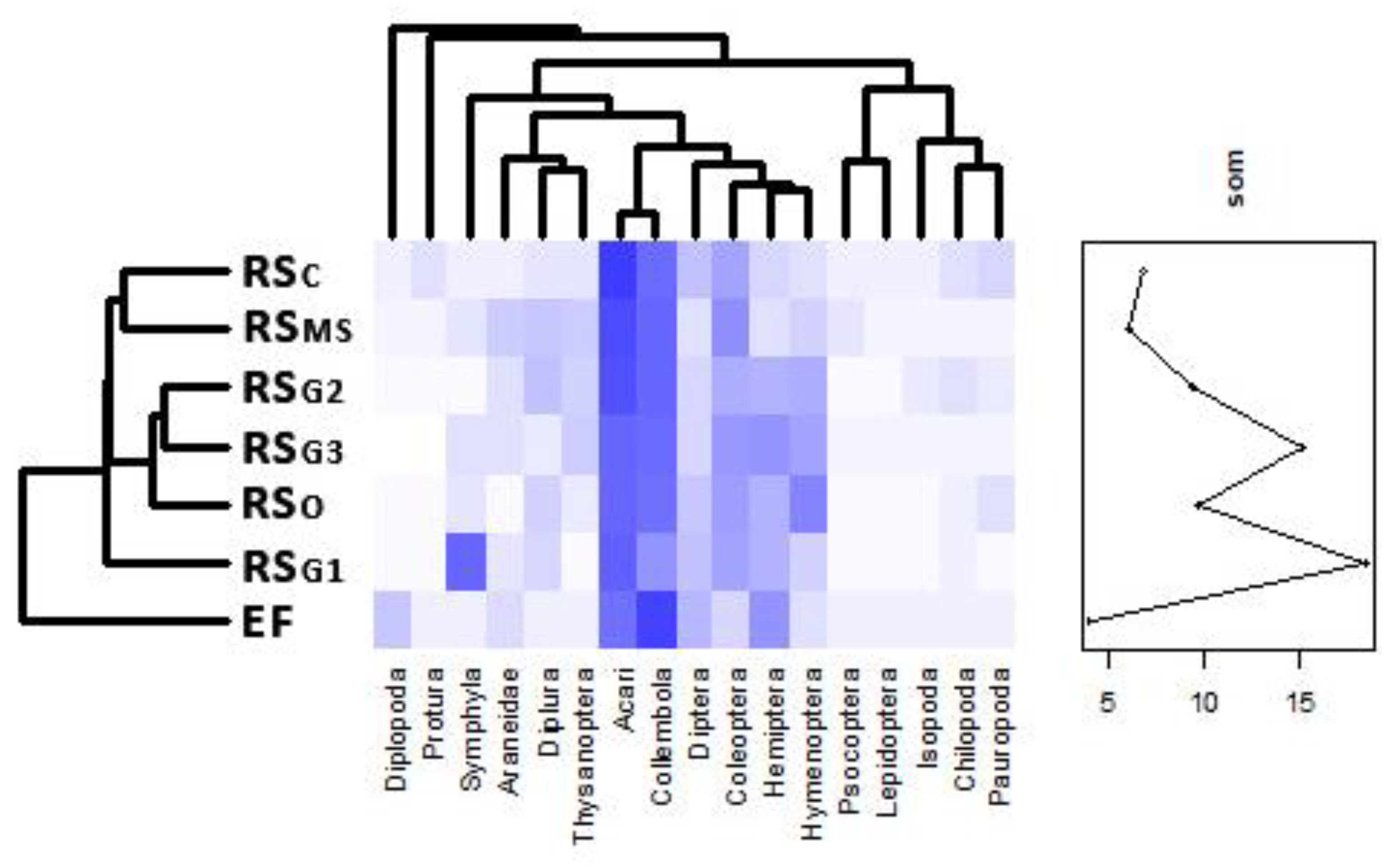
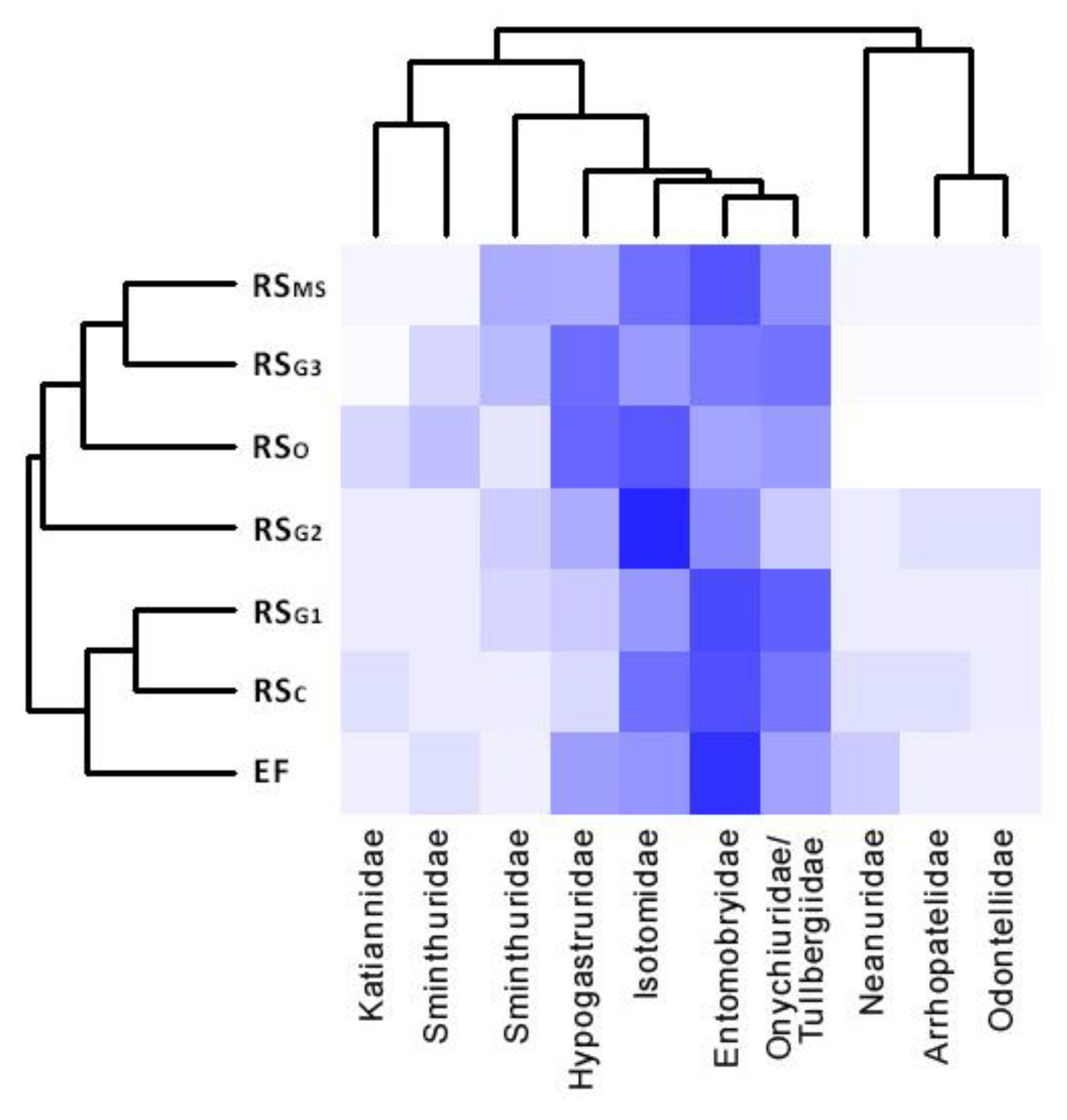
3.4. Soil Microarthropod Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boccaletti, M.; Elter, P.; Guazzone, G. Plate Tectonic Models for the Development of the Western Alps and Northern Apennines. Nat. Phys. Sci. 1971, 234, 108–111. [Google Scholar] [CrossRef]
- Kligfield, R. The northern Apennines as a collisional orogen. Am. J. Sci. 1979, 279, 676–691. [Google Scholar] [CrossRef]
- Vai, G.B.; Martini, P. Anatomy of an Orogen: The Apennines and Adjacent Mediterranean Basins; Kluwer Academic Publishers: Tranbjerg, Denmark, 2001; ISBN 9780412750403. [Google Scholar]
- Molli, G. Northern Apennine–Corsica orogenic system: An updated overview. Geol. Soc. Lond. Spec. Publ. 2008, 298, 413–442. [Google Scholar] [CrossRef]
- Crozier, M.J. Landslide geomorphology: An argument for recognition, with examples from New Zealand. Geomorphology 2010, 120, 3–15. [Google Scholar] [CrossRef]
- Carlini, M.; Chelli, A.; Vescovi, P.; Artoni, A.; Clemenzi, L.; Tellini, C.; Torelli, L. Tectonic control on the development and distribution of large landslides in the Northern Apennines (Italy). Geomorphology 2016, 253, 425–437. [Google Scholar] [CrossRef]
- Perrone, A.; Iannuzzi, A.; Lapenna, V.; Lorenzo, P.; Piscitelli, S.; Rizzo, E.; Sdao, F. High resolution electrical imaging of the Varco d’Izzo earthflow (southern Italy). J. Appl. Geophys. 2004, 56, 17–29. [Google Scholar] [CrossRef]
- Bertolini, G.; Corsini, A.; Tellini, C. Fingerprints of Large-Scale Landslides in the Landscape of the Emilia Apennines. In Landscapes and Landforms of Italy; Soldati, M., Marchetti, M., Eds.; Springer International Publishing: Berlin, Germany, 2017; pp. 215–224. [Google Scholar] [CrossRef]
- Carlini, M.; Chelli, A.; Francese, R.; Giacomelli, S.; Giorgi, M.; Quagliarini, A.; Carpena, A.; Tellini, C. Landslides types controlled by tectonics-induced evolution of valley slopes (Northern Apennines, Italy). Landslides 2018, 15, 283–296. [Google Scholar] [CrossRef]
- Antolini, G.; Pavan, V.; Tomozeiu, R.M.V. Atlante Climatico dell’Emilia-Romagna 1961–2015; ARPAe: Bologna, Italy, 2017; ISBN 9788887854442. [Google Scholar]
- Jongman, B.; Hochrainer-Stigler, S.; Feyen, L.; Aerts, J.C.J.H.; Mechler, R.; Botzen, W.J.W.; Bouwer, L.M.; Pflug, G.; Rojas, R.; Ward, P.J. Increasing stress on disaster-risk finance due to large floods. Nat. Clim. Chang. 2014, 4, 264–268. [Google Scholar] [CrossRef]
- Palis, E.; Lebourg, T.; Vidal, M.; Levy, C.; Tric, E.; Hernandez, M. Multiyear time-lapse ERT to study short- and long-term landslide hydrological dynamics. Landslides 2017, 14, 1333–1343. [Google Scholar] [CrossRef]
- Bièvre, G.; Jongmans, D.; Goutaland, D.; Pathier, E.; Zumbo, V. Geophysical characterization of the lithological control on the kinematic pattern in a large clayey landslide (Avignonet, French Alps). Landslides 2016, 13, 423–436. [Google Scholar] [CrossRef]
- Segoni, S.; Piciullo, L.; Gariano, S.L. A review of the recent literature on rainfall thresholds for landslide occurrence. Landslides 2018, 15, 1483–1501. [Google Scholar] [CrossRef]
- Lissak, C.; Maquaire, O.; Malet, J.P.; Bitri, A.; Samyn, K.; Grandjean, G.; Bourdeau, C.; Reiffsteck, P.; Davidson, R. Airborne and ground-based data sources for characterizing the morphostructure of a coastal landslide. Geomorphology 2014, 217, 140–151. [Google Scholar] [CrossRef]
- Geertsema, M.; Pojar, J.J. Influence of landslides on biophysical diversity—A perspective from British Columbia. Geomorphology 2007, 89, 55–69. [Google Scholar] [CrossRef]
- Geertsema, M.; Highland, L.; Vaugeouis, L. Environmental Impact of Landslides. In Landslides—Disaster Risk Reduction; Springer: Berlin/Heidelberg, Germany, 2009; pp. 589–607. ISBN 9783540699705. [Google Scholar] [CrossRef]
- Pimentel, D. Soil erosion: A food and environmental threat. Environ. Dev. Sustain. 2006, 8, 119–137. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Smith, R.B.; Commandeur, P.R.; Ryan, M.W. Soils, Vegetation, and Forest Growth on Landslides and Surrounding Logged and Old-Growth Areas on the Queen Charlotte Islands; BC Land Management Report; Ministry of Forests: Victoria, BC, Canada, 1986; Volume 41, p. 95.
- Vescovi, P. Note Illustrative Della Carta Geologica D’italia Alla Scala 1:50.000. Foglio 216 Borgo Val di Taro; Servizio Geologico d’Italia-Regione Emilia Romagna: Roma, Italy, 2002; p. 115. [Google Scholar]
- Cruden, D.M.; Varnes, D.J. Landslide Types and Processes, Special Report. In Landslides: Investigation and Mitigation; Turner, A.K., Shuster, R.L., Eds.; National Academy Press: Washington, DC, USA, 1996; Volume 247, pp. 36–75. [Google Scholar]
- Cartografia Interattiva e Banche Dati. Available online: Ambiente.regione.emilia-romagna.it/it/geologia/cartografia/webgis-banchedati (accessed on 5 August 2019).
- Morin, R.H.; Carleton, G.B.; Poirier, S. Fractured-aquifer hydrogeology from geophysical logs; the passaic formation, New Jersey. Ground Water 1997, 35, 328–338. [Google Scholar] [CrossRef]
- Cook, P.G.; Love, A.J.; Dighton, J.C. Inferring ground water flow in fractured rock from dissolved radon. Ground Water 1999, 37, 606–610. [Google Scholar] [CrossRef]
- Petrella, E.; Naclerio, G.; Falasca, A.; Bucci, A.; Capuano, P.; De Felice, V.; Celico, F. Non permanent shallow halocline in a fractured carbonate aquifer, southern Italy. J. Hydrol. 2009, 373, 267–272. [Google Scholar] [CrossRef]
- Aquino, D.; Petrella, E.; Florio, M.; Celico, P.; Celico, F. Complex hydraulic interactions between compartmentalized carbonate aquifers and heterogeneous siliciclastic successions: A case study in southern Italy. Hydrol. Process. 2015, 29, 4252–4263. [Google Scholar] [CrossRef]
- FAO. Guidelines for Profile Description, 3rd ed.; Soil Resources, Management and Conservation Service, Land and Water Development Division; FAO: Rome, Italy, 1990. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall, Inc.: Englewood Cliffs, NJ, USA, 1958; 498 S. DM 39.40. Zeitschrift für Pflanzenernährung Düngung Bodenkd. 2007, 85, 251–252. [Google Scholar] [CrossRef]
- Ball, D.F. Loss-On-Ignition As An Estimate Of Organic Matter And Organic Carbon In N -Calcareous Soils. J. Soil Sci. 1964, 15, 84–92. [Google Scholar] [CrossRef]
- The Taxonomicon. Available online: http://taxonomicon.taxonomy.nl/ (accessed on 5 August 2019).
- Bachelier, P.G. La vie Animale Dans les sols I. -Déterminisme de la Faune des Sols; ORSTOM: Paris, France, 1963. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P.; Borcard, D.; Gillet, F.; Legendre, P. Association Measures and Matrices. In Numerical Ecology with R; Springer: New York, NY, USA, 2011; pp. 31–51. ISBN 978-1-4419-7975-9. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 23 February 2019).
- Petrella, E.; Celico, F. Mixing of water in a carbonate aquifer, southern Italy, analysed through stable isotope investigations. Int. J. Speleol. 2013, 42, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Wilcke, W.; Valladarez, H.; Stoyan, R.; Yasin, S.; Valarezo, C.; Zech, W. Soil properties on a chronosequence of landslides in montane rain forest, Ecuador. CATENA 2003, 53, 79–95. [Google Scholar] [CrossRef]
- Chibsa, T.; Ta, A.A. Assessment of Soil Organic Matter under Four Land Use Systems in the Major Soils of Bale Highlands, South East Ethiopia b. Factors Affecting Soil Organic Matter Distribution. World Appl. Sci. J. 2009, 6, 1506–1512. [Google Scholar]
- Walker, L.R.; Velázquez, E.; Shiels, A.B. Applying lessons from ecological succession to the restoration of landslides. Plant Soil 2009, 324, 157–168. [Google Scholar] [CrossRef]
- Menta, C.; Leoni, A.; Gardi, C.; Conti, F. Are grasslands important habitats for soil microarthropod conservation? Biodivers. Conserv. 2011, 20, 1073–1087. [Google Scholar] [CrossRef]
- Parisi, V.; Menta, C.; Gardi, C.; Jacomini, C.; Mozzanica, E. Microarthropod Communities as a Tool to Assess Soil Quality and Biodiversity: A new Approach in Italy. Agric. Ecosyst. Environ. 2005, 105, 323–333. [Google Scholar] [CrossRef]
- Parisi, V.; Menta, C. Microarthropods of the soil: Convergence phenomena and evaluation of soil quality using QBS-ar and QBS-c. Fresenius Environ. Bull. 2008, 17, 1170–1174. [Google Scholar]
- Van Vliet, P.C.J.; Hendrix, P.F. Role of fauna in soil physical processes. In Soil Biological Fertility: A Key to Sustainable Land Use in Agriculture; Springer: Dordrecht, The Netherlands, 2007; pp. 61–80. ISBN 9781402066184. [Google Scholar] [CrossRef]
- Rusek, J. Soil microstructures-contributions on specific soil organisms. Quest. Entomol. 1985, 21, 497–514. [Google Scholar]
- Schatz, H.; Behan-Pelletier, V. Global diversity of oribatids (Oribatida: Acari: Arachnida). Hydrobiologia 2008, 595, 323–328. [Google Scholar] [CrossRef]
- Maraun, M.; Scheu, S. The Structure of Oribatid Mite Communities (Acari, Oribatida): Patterns, Mechanisms and Implications for Future Research. Ecography 2000, 23, 374–383. [Google Scholar] [CrossRef]
- Gergocs, V.; Hufnagel, L. Application of Oribatid Mites as Indicators (Review). Appl. Ecol. Environ. Res. 2009, 7, 79–98. [Google Scholar] [CrossRef]
- Lavelle, P. (Patrick); Spain, A.V. Soil organisms. In Soil ecology; Springer Science & Business Media, Ed.; Springer: Dordrecht, The Netherlands, 2007; p. 654. ISBN 9780306481628. [Google Scholar]
- Brown, V.K.; Gange, A.C. Insect Herbivory Insect Below Ground. Adv. Ecol. Res. 1990, 20, 1–58. [Google Scholar]
- Gabet, E.J.; Reichman, O.J.; Seabloom, E.W. The effects of bioturbation on soil processes and sediment transport. Annu. Rev. Earth Planet. Sci. 2003, 31, 249–273. [Google Scholar] [CrossRef]
- Frouz, J. Use of soil dwelling Diptera (Insecta, Diptera) as bioindicators: A review of ecological requirements and response to disturbance. Agric. Ecosyst. Environ. 1999, 74, 167–186. [Google Scholar] [CrossRef]
- Edwards, C.A. The ecology of Symphyla. Entomol. Exp. Appl. 1958, 1, 308–319. [Google Scholar] [CrossRef]
- Petrella, E.; Bucci, A.; Ogata, K.; Zanini, A.; Naclerio, G.; Chelli, A.; Francese, R.; Boschetti, T.; Pittalis, D.; Celico, F. Hydrodynamics in Evaporate-Bearing Fine-Grained Successions Investigated through an Interdisciplinary Approach: A Test Study in Southern Italy—Hydrogeological Behaviour of Heterogeneous Low-Permeability Media. Geofluids 2018, 2018. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Code | Movement Type | Land Use | Coordinates (UTM) |
|---|---|---|---|
| EF | Earth flow | Eroded zone belonging to the active earth flow area | 32N 573645 4938131 |
| RSC | Rotational slide | Wheat cultivated field | 32N 573634 4938510 |
| RSG1 | Rotational slide | Permanent grassland | 32N 573628 4938442 |
| RSG2 | Rotational slide | Permanent grassland | 32N 573732 4938321 |
| RSG3 | Rotational slide | Permanent grassland | 32N 573764 4938410 |
| RSMS | Rotational slide | Main scarp with scarce vegetation cover | 32N 573645 4938357 |
| RSO | Rotational slide | Overgrown field | 32N 573641 4938486 |
| Faunal Group | EF | RSc | RSG1 | RSG2 | RSG3 | RSMS | RSO |
|---|---|---|---|---|---|---|---|
| Acari | 311 ± 79 | 7268 ± 5867 | 7844 ± 447 | 27,588 ± 2654 | 4681 ± 313 | 6008 ± 3273 | 4897 ± 1345 |
| Araneidae | 14 ± 7 | 21 ± 0 | 42 ± 12 | 42 ± 12 | 64 ± 37 | - | |
| Chilopoda | - | 14 ± 7 | 11 ± 6 | 42 ± 25 | 11 ± 6 | - | 7 ± 7 |
| Coleoptera | 14 ± 7 | 255 ± 131 | 573 ± 123 | 499 ± 67 | 796 ± 190 | 594 ± 294 | 566 ± 209 |
| Larvae | - | 255 ± 161 | 541 ± 244 | 403 ± 74 | 786 ± 196 | 531 ± 305 | 559 ± 203 |
| Collembola | 870 ± 492 | 1755 ± 494 | 1040 ± 123 | 10,870 ± 650 | 3917 ± 1122 | 2420 ± 735 | 3276 ± 1769 |
| Diplopoda | 28 ± 19 | - | - | - | - | - | - |
| Diplura | - | 7 ± 7 | 53 ± 18 | 202 ± 80 | 21 ± 12 | 74 ± 18 | 64 ± 25 |
| Diptera | 50 ± 7 | 78 ± 58 | 159 ± 43 | 74 ± 18 | 74 ± 6 | 21 ± 7 | 120 ± 57 |
| Larvae | 14 ± 7 | 64 ± 44 | 42 ± 21 | 64 ± 25 | 11 ± 6 | 11 ± 7 | 64 ± 12 |
| Hemiptera | 142 ± 131 | 28 ± 14 | 276 ± 86 | 393 ± 153 | 902 ± 337 | 21 ± 12 | 248 ± 146 |
| Hymenoptera | 7 ± 7 | - | 85 ± 37 | 510 ± 270 | 510 ± 221 | 42 ± 25 | 1720 ± 1481 |
| Isopoda | - | - | - | 21 ± 12 | 11 ± 6 | - | - |
| Lepidoptera | - | - | - | - | 11 ± 6 | - | - |
| Larvae | - | - | - | - | 11 ± 6 | - | - |
| Pauropoda | - | 28 ± 19 | - | 21 ± 12 | 11 ± 6 | - | 28 ± 14 |
| Protura | - | 14 ± 14 | - | - | - | - | - |
| Psocoptera | - | - | - | - | 11 ± 6 | 11 ± 6 | - |
| Symphyla | - | - | 6496 ± 2415 | - | 42 ± 12 | 11 ± 6 | 21 ± 12 |
| Thysanoptera | - | 7 ± 7 | - | 117 ± 6 | 106 ± 61 | 53 ± 31 | 14 ± 7 |
| Number of groups | 8 ± 1 | 10 ± 0 | 10 ± 0 | 11 ± 1 | 13 ± 1 | 10 ± 1 | 10 ± 0 |
| Total abundance | 1437 ± 709 | 9469 ± 5699 | 16,559 ± 2674 | 40,379 ± 3711 | 11,146 ± 1243 | 9320 ± 4380 | 10,962 ± 3910 |
| Acari/Collembola | 2 ± 1 | 6 ± 5 | 8 ± 1 | 3 ± 0 | 1 ± 0 | 2 ± 1 | 2 ± 1 |
| Collembola Family | EF | RSC | RSG1 | RSG2 | RSG3 | RSMS | RSO |
|---|---|---|---|---|---|---|---|
| Arrhopalitidae | - | 7 ± 7 | - | 11 ± 6 | - | - | - |
| Entomobryidae | 623 ± 429 | 934 ± 532 | 563 ± 67 | 393 ± 80 | 955 ± 110 | 1306 ± 215 | 205 ± 51 |
| Hypogastruridae | 71 ± 51 | 14 ± 7 | 21 ± 12 | 138 ± 18 | 1337 ± 735 | 106 ± 61 | 1076 ± 416 |
| Isotomidae | 85 ± 65 | 425 ± 170 | 96 ± 31 | 10,243 ± 766 | 329 ± 116 | 616 ± 282 | 1592 ± 1392 |
| Neanuridae | 21 ± 12 | 7 ± 7 | - | - | - | - | - |
| Onychiuridae/Tullbergiidae | 64 ± 64 | 361 ± 165 | 350 ± 67 | 42 ± 12 | 1136 ± 313 | 265 ± 104 | 255 ± 44 |
| Odontellidae | - | - | - | 11 ± 6 | - | - | - |
| Katiannidae | - | 7 ± 7 | - | - | - | - | 42 ± 21 |
| Sminthuridae | - | - | 11 ± 6 | 32 ± 6 | 117 ± 55 | 127 ± 74 | 21 ± 12 |
| Sminthurididae | 7 ± 7 | - | - | - | 42 ± 25 | - | 85 ± 56 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Remelli, S.; Petrella, E.; Chelli, A.; Conti, F.D.; Lozano Fondón, C.; Celico, F.; Francese, R.; Menta, C. Hydrodynamic and Soil Biodiversity Characterization in an Active Landslide. Water 2019, 11, 1882. https://doi.org/10.3390/w11091882
Remelli S, Petrella E, Chelli A, Conti FD, Lozano Fondón C, Celico F, Francese R, Menta C. Hydrodynamic and Soil Biodiversity Characterization in an Active Landslide. Water. 2019; 11(9):1882. https://doi.org/10.3390/w11091882
Chicago/Turabian StyleRemelli, Sara, Emma Petrella, Alessandro Chelli, Federica Delia Conti, Carlos Lozano Fondón, Fulvio Celico, Roberto Francese, and Cristina Menta. 2019. "Hydrodynamic and Soil Biodiversity Characterization in an Active Landslide" Water 11, no. 9: 1882. https://doi.org/10.3390/w11091882