1. Introduction
The reasonable use of natural resources requires a balance to be maintained between the improvement of the quality of life and fast economic growth on the one hand, and the improvement of the natural environment and the protection of its resources on the other. Overuse of environmental resources through intensive agriculture, industry, spatial development of cities and villages, architecture, transport, infrastructure development and inappropriate land management can contribute greatly towards the destabilization of the environment, especially in urbanized ecosystems [
1].
Urban bottom sediments are naturally formed in water reservoirs during the sedimentation of mineral and organic suspensions, as well as components precipitated from water. Due to the rapid rate of accumulation of bottom sediments, they should be periodically removed in order to enhance reservoir capacity and to improve its functional value [
2,
3]. One example of a reservoir that is also an urban area sedimentation pond is the Sokołówka sequential biofiltration system (SSBS). The SSBS was constructed in the upper section of the Sokołówka River with the aim of removing various pollutants, such as sediments, suspended solids, particulate pollutants, petroleum hydrocarbons, heavy metals and nutrients from stormwater runoff. To do so, it employs a system of sedimentation and filtration mechanisms. The system comprises of three different zones to improve efficiency: A hydrodynamically intensified sedimentation zone, an intensive biogeochemical zone and an intensive biofiltration zone [
4].
The composition of the bottom sediments depends on many factors, such as the way the catchment is used, its natural conditions and on the type and amount of contaminants reaching the surface waters. In many locations, bottom sediments include the material brought along with industrial and municipal sewage, as well as surface runoff from urbanized, industrial and agricultural areas [
5]. The most common contaminants in bottom sediments include heavy metals and inorganic compounds such as silicates and aluminates, as well as pathogenic and other microbial pollutants; these are accompanied by a number of toxic organic pollutants, such as persistent organic pollutants (POPs), which are characterized by high persistence and great potential for bioaccumulation, biomagnification and toxicity; e.g., polychlorinated biphenyls (PCBs), polychlorinated dibenzo-
p-dioxin (PCDD), polychlorinated dibenzofurans (PCDFs) and polycyclic aromatic hydrocarbons (PAH) [
6,
7,
8,
9,
10].
On the other hand, bottom sediments may also act as fertilizers: Following decomposition, they serve as a rich source of essential organic and mineral matter for plants, including nitrogen and phosphorus compounds, that have important agricultural applications [
9,
11,
12]. In addition, organic fertilization supplements the organic carbon and nitrogen stocks in the soil to a greater degree than mineral fertilization [
13,
14]. Such supplementation improves soil carbon sequestration and is particularly important in the face of climate change. Soils with higher levels of organic matter also demonstrate better structure, greater water holding capacity and increased soil nutrient availability; they also have greater microbial biomass, as well as a more diverse microbial community structure and greater biodiversity in general [
15,
16]. Organic fertilization can be particularly useful in cities, with urban bottom sediments offering the potential for the reclamation of degraded land, communication areas and recreational areas, such as flower beds and greenery, where ornamental plants and lawns are mainly grown.
It is possible to enhance the fertilizer properties of urban sediments while also negating their toxicity through the use of remediation techniques. One such technique is phytoremediation. It uses the ability and metabolic activity of selected plant species and their symbiotic microorganisms to reclaim a contaminated environment [
17,
18,
19,
20,
21]. The very presence of a root system allows the immobilization of the soil and pollutants in a contaminated substrate, inhibiting erosion by air and water while also preventing the movement of contaminants to deeper layers of the soil profile. At this stage, such immobilization is particularly important in the presence of contaminants characterized by poor water solubility and low biodegradability, including POPs such as PCBs, PCDD, PCDFs or PAH. Although such remediation typically takes place via uptake by plant roots, it also occurs outside the roots through the participation of enzymes secreted by the plants into the rhizosphere, which stimulate the development of microorganisms responsible for detoxification. This use of plant roots to maintain a favorable growth environment for the contaminant-degrading rhizosphere microorganisms in soil is named rhizodegradation [
22,
23]. The rhizosphere, the area comprising the soil surrounding the root and the root surface itself, is characterized by intense microbiological activity. The biological activity taking place in this zone is one of the factors conditioning the growth of plants and their resistance to pollution [
24]. However, the development of the root zone not only increases the total number and diversity of microorganisms, the plants also change the physicochemical properties of the local soil, improve the water and air ratios and maintain the correct, crumbly soil structure; they also modulate the pH, organic carbon content and mineral content of the soil. These factors contribute to the creation of favorable conditions for the course of biological processes, thereby stimulating bioremediation.
Many examples of relationships between plants and microorganisms that show potential applications in phytoremediation techniques have been observed, some of which use selected plant growth-promoting bacteria (PGPB). However, despite being a potentially valuable method for managing sediment-contaminated soils, there is currently a lack of evidence surrounding the use of bacterial inoculants in remediation, with most previous studies on the topic focusing on contaminated soils [
25]. One of the bacterial strains used in recent research is
Massilia niastensis p87, which was isolated from the rhizosphere of
Festuca rubra growing on mine tailings with elevated concentrations of Cd, Pb and Zn. Strain p87 is a Cd/Zn-resistant bacterium that is an indole-3-acetic acid-producer, and was previously found to increase the biomass of both
Salix viminalis and
Festuca arundinacea growing in Cd/Zn-enriched Hoagland solutions [
26]. Another bacterial strain is
Streptomyces costaricanus RP92, which was isolated from the rhizosphere of
Cytisus striatus (Hill) Rothm. growing in hexachlorocyclohexane (HCH)-contaminated soil; it has been characterized as an IAA and siderophore-producer [
27]. Strain RP92 was previously shown to stimulate the growth of
Lupinus luteus in diesel-contaminated soils, as well as to improve the dissipation of diesel range organics [
28].
Tagetes patula L., commonly called French marigold, is a dicotyledonous annual plant species from the Asteraceae family, native from Mexico to Argentina. It is a well-known ornamental and medicinal plant, known to be a source of various secondary compounds including triterpene, thiophene and steroids. These compounds may be variably excreted by the plant in response to altering environmental conditions and have been shown to have antimicrobial, insecticidal and nematicidal effects [
29,
30]. It has been reported that
T. patula is a very effective plant in benzo[
a]pyrene remediation and offers substantial potential for soil metal remediation [
31,
32].
Festuca arundinacea (Schreb.), popularly known as tall fescue, is a monocotyledon species of perennial grass belonging to the Poaceae family. It is very resistant to adverse climatic conditions. Due to its low habitat requirements, it is used on areas devastated by industry and is a popular choice for phytoremediation [
33,
34].
In addition to certain physical characteristics such as fast growth with high biomass production, and a deep rooting system that is easily harvestable, the plants used for phytoremediation should also possess many features that will be conducive to survival in the difficult conditions of a contaminated environment, e.g., large tolerance for the uptake and accumulation of pollutants in their tissues. The potential presence of toxic substances in bottom sediments may act as an environmental stress factor and may have an adverse effect on plant growth and development by disturbing plant internal homeostasis. On the other hand, inoculation of the soil with selected plant growth-promoting bacterial strains may not only increase the effectiveness of the phytoremediation process but also contribute to alleviating the effects of stress factors on plant organisms.
The purpose of the present work was (1) to examine the response of selected plant species to soil application of bottom sediments from the urban reservoir, and to (2) determine whether the addition of selected bacterial strains supports the development and condition of plants grown on the soil without the addition of sediments; in addition, it examines (3) whether the addition of these bacterial strains influences the reaction of plants to the presence of bottom sediments in the substrate. The yield of the aboveground biomass and physiological status of the two species of investigated plants is determined by measuring total soluble protein and total chlorophyll content and chlorophyll a/b ratio. In addition, to determine the influence of the contaminants on the redox equilibrium of the plant, lipid peroxide content was measured as thiobarbituric acid reactive substances (TBARS), as well as non-enzymatic antioxidants such as α-tocopherol and total phenolic compound content (TPC). The anthocyanins content was also analyzed as a marker for the occurrence of environmental stress.
3. Results
The soil and sediment samples were analyzed for total concentration of trace elements, sixteen PAH compounds and seventeen toxic congeners of PCDD/PCDF. As presented in our previous work, the sediments were moderately contaminated with zinc and heavily contaminated with PAH, PCDD and PCDFs [
35].
The addition of bottom sediments and the inoculation of bacterial strains were found to have different influences on the plants depending on the species.
3.1. Plant Morphology and Dry Biomass
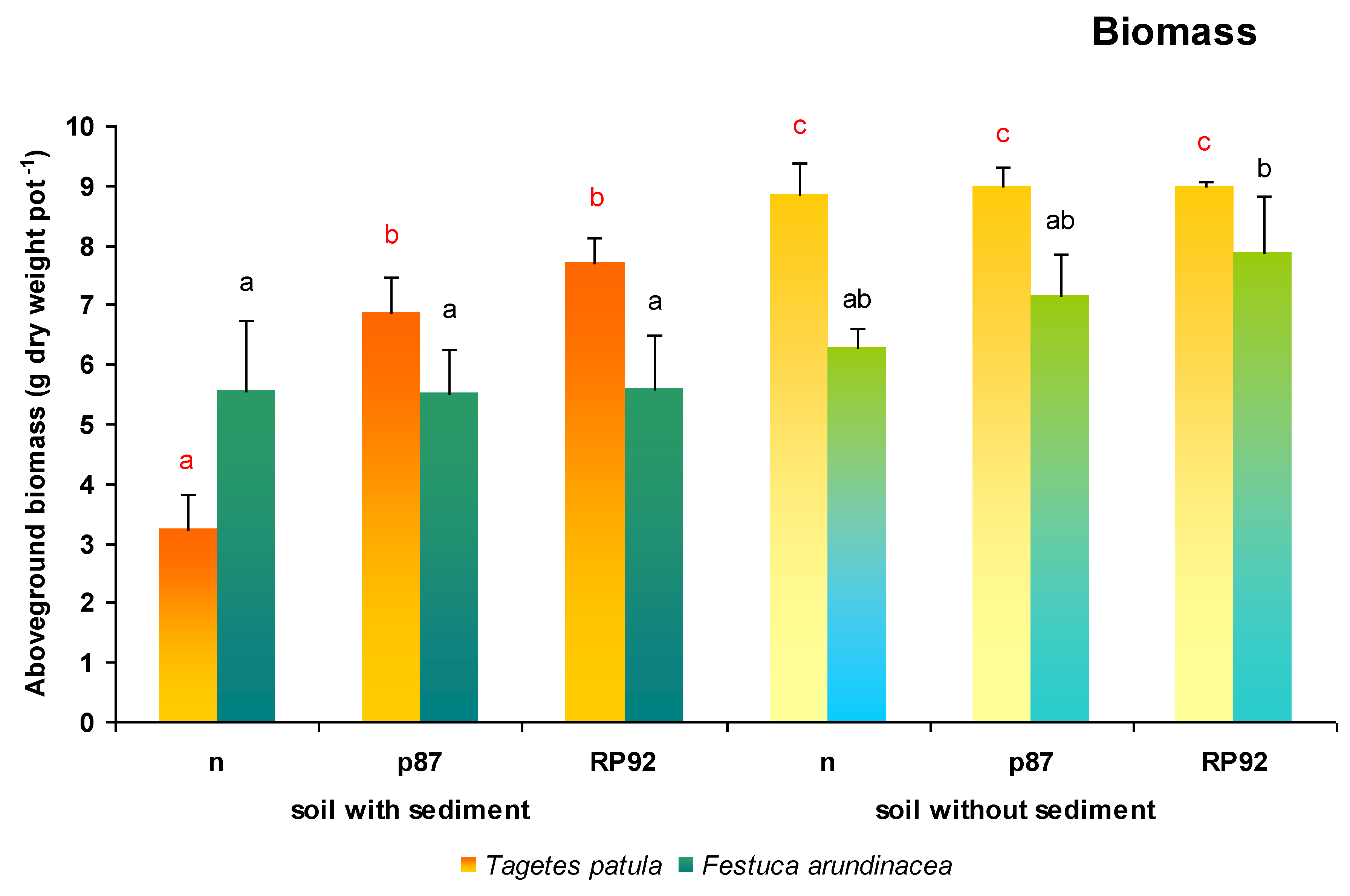
T. patula plants growing on sediment-amended soil were characterized by very poor biomass growth and inhibited development (flower buds did not develop) compared to the other variants i.e., those inoculated with bacterial strains p87 and RP92, as well as those grown on non-amended soil (
Figure 1). The non-inoculated plants grown on the amended soil had characteristic reddish-purple discolorations and slight necrosis at the edge of the leaves. However, the plants growing on amended soil with bacterial inoculation were well developed; those with p87 displayed a large number of young shoots in the leaves at a later stage of development, and those inoculated with RP92 were characterized by an intense green color. In addition, the leaves of plants grown on sediment-amended soil and inoculated with p87 or RP92 were in good general condition, with very few being wrinkled, or folded with incorrect structure.
T. patula grown on non-amended soil demonstrated proper development, and bacterial inoculation had no effect on plant morphology. In
F. arundinacea, neither inoculation with bacterial strains nor the presence of sediment appeared to have any significant influence on morphology.
In
T. patula plants, both the presence of sediments in the soil (F = 122.756; df = 1, 18;
p = 0.0000) and the type of inoculation had an effect on the biomass yield (F = 27.145; df = 2, 18;
p = 0.0000; ANOVA II;
Figure 2). The addition of urban sediments significantly reduced the biomass yield of
T. patula plants. If bacterial inoculates were also added to the soil with urban sediments, the yield increased to 213% (p87) and to 238% (RP92) compared to plants growing on non-inoculated soil. The yield of
T. patula plants growing on soil without sediment was 274% of that of plants growing on soil with sediment. The use of bacterial strains on soil without sediment also increased the yield to 131% (p87) and to 117% (RP92) compared to plants growing on soil with sediment and inoculated with strains p87 and RP92, respectively. Inoculation was found to have no effect on biomass in plants growing on soil without sediments.
The biomass yield in F. arundinacea plants depended only on the presence of sediment in soil (F = 7.4795; df = 1, 18; p = 0.0136). F. arundinacea plants did not show differences in biomass yield, either after using urban sediments or inoculation with bacterial strains. The only exception was observed for plants growing on soil without sediment and inoculated with RP92 strain, which demonstrated 41%–42% greater biomass than plants growing on soil with sediments.
3.2. Total Soluble Protein Content
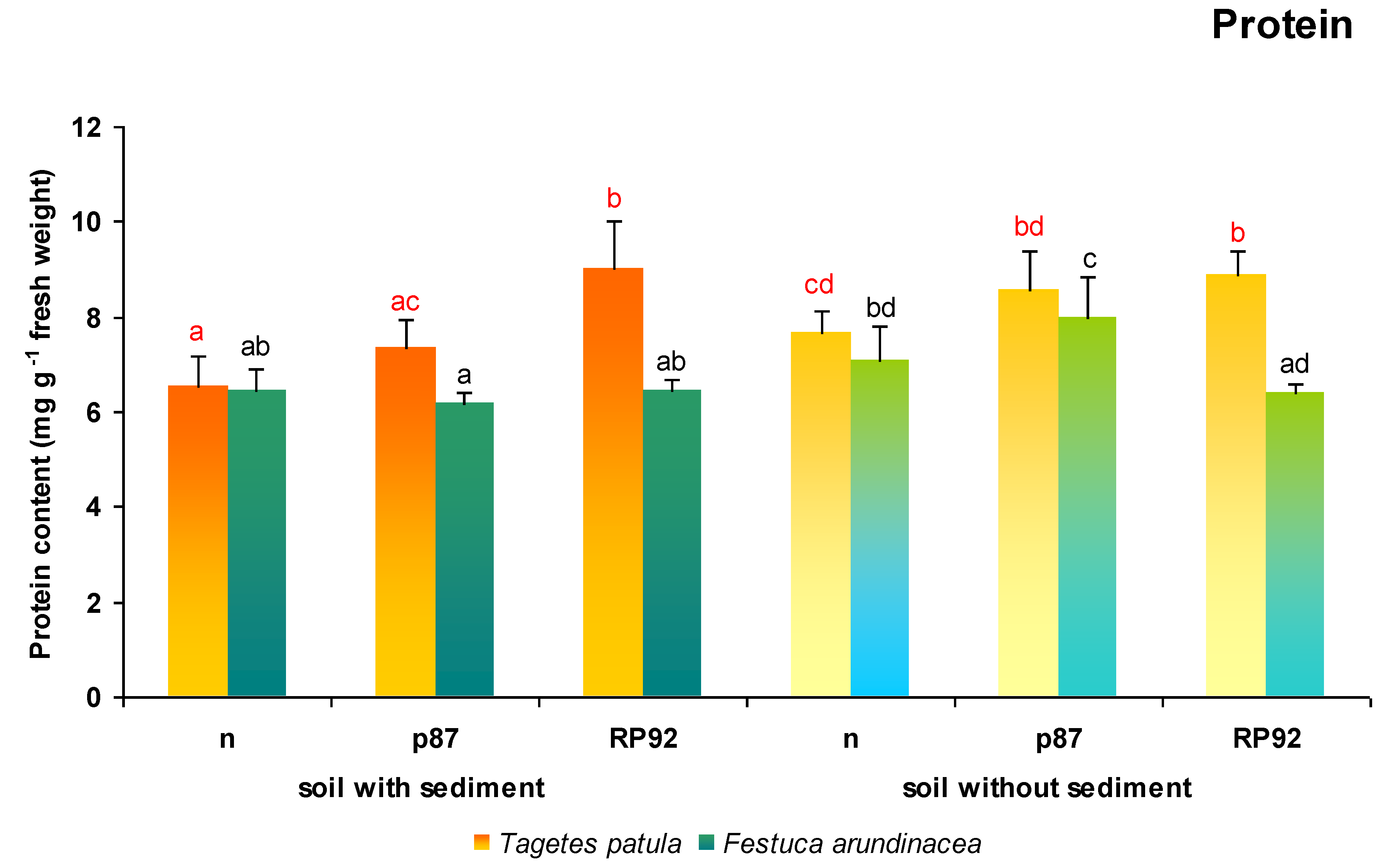
Changes in soluble protein content in
T. patula leaf tissues were influenced by the presence of sediment in soil and application of inoculation (
Figure 3). ANOVA II showed that the presence of sediment (F = 6.98; df = 1, 18;
p = 0.0166) and bacterial inoculation (F = 15.102; df = 2, 18;
p = 0.000141) both influenced soluble protein content. The Duncan test revealed that the use of bottom sediments caused a decrease in soluble protein content compared to plants growing on soil without sediments: It was 15% lower than for plants growing on non-inoculated soil and 14% lower than in soil inoculated with the p87 strain.
The use of the RP92 strain significantly increased the soluble protein content in T. patula leaves compared to plants growing on non-inoculated soil, both after the application of sediment (139%) and on the soil without sediment (116%). In contrast, no such dependence was found after the inoculation with the p87 strain. Moreover, the soluble protein content of the plants grown on the soil containing sediments was found to be significantly higher after inoculation with the RP92 strain than the p87 strain (123%). The application of p87 caused a significant increase in protein content, but only among plants growing on the non-enriched substrate, compared to those growing on non-inoculated soil enriched with sediments (131%).
In the case of F. arundinacea, ANOVA II showed that only the use of sediments had a significant effect on the soluble protein content (F = 14.737; df = 1, 18; p = 0.0012). The Duncan test showed that the use of the p87 strain on soil to which no sediments were added significantly increased the soluble protein content in leaf tissues compared to all other variants (from 112% to 129%). Interestingly, while the use of the p87 strain with the sediments reduced the total soluble protein content in plant tissues to 87%, inoculation without the simultaneous use of sediments increased the protein content to 112%: both cases compared to the non-inoculated variants.
3.3. Total Chlorophyll Content
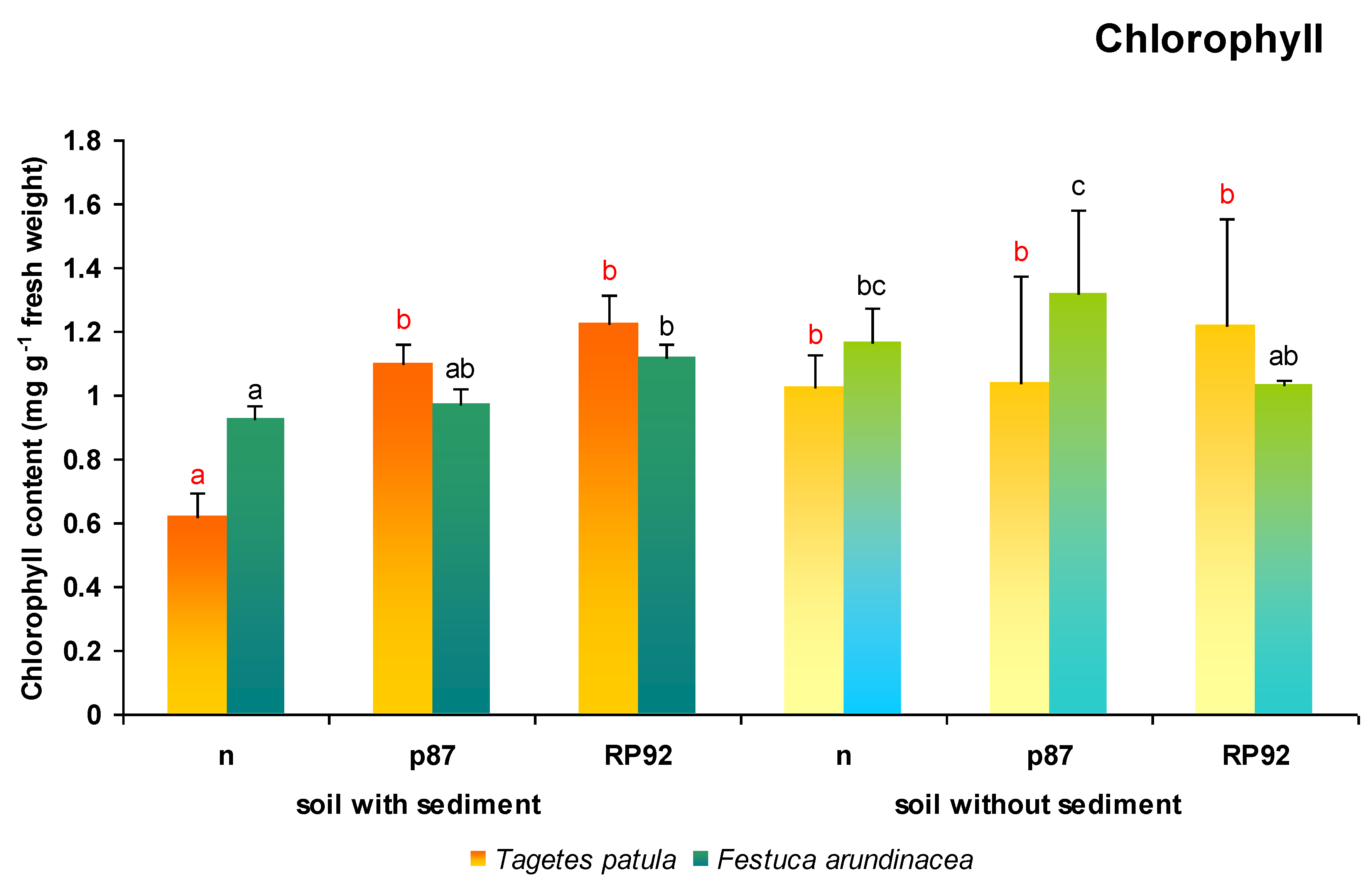
The presence of sediment was not found to influence total chlorophyll content in
T. patula leaves (ANOVA II). In contrast, the type of inoculation dependent changes in the value of this parameter was observed (F = 7.901; df = 2, 18;
p = 0.0034). The Duncan post hoc test indicated that the total chlorophyll content of
T. patula plants growing in sediment-enriched soil without inoculum was significantly lower than in all other experimental variants (
Figure 4).
The total chlorophyll content in F. arundinacea was dependent on the presence of sediments in the soil (F = 11.300; df = 1, 18; p = 0.0035); however, it was not dependent on the type of inoculation used. The Duncan’s post hoc test revealed that plants growing on non-inoculated soil presented higher total chlorophyll content when the soil was not enriched with sediments (126%). Moreover, for plants growing on soil with sediments, RP92 inoculation increased the total chlorophyll content (121%) compared to plants growing on non-inoculated soil with sediments. The highest total chlorophyll content was found in plants growing on soil without sediments inoculated with the p87 strain, with the content ranging from 117% to 142% compared to the other variants; this value was also higher than in plants growing on non-amended soil inoculated with strain RP92.
3.4. Chlorophyll a/b Ratio
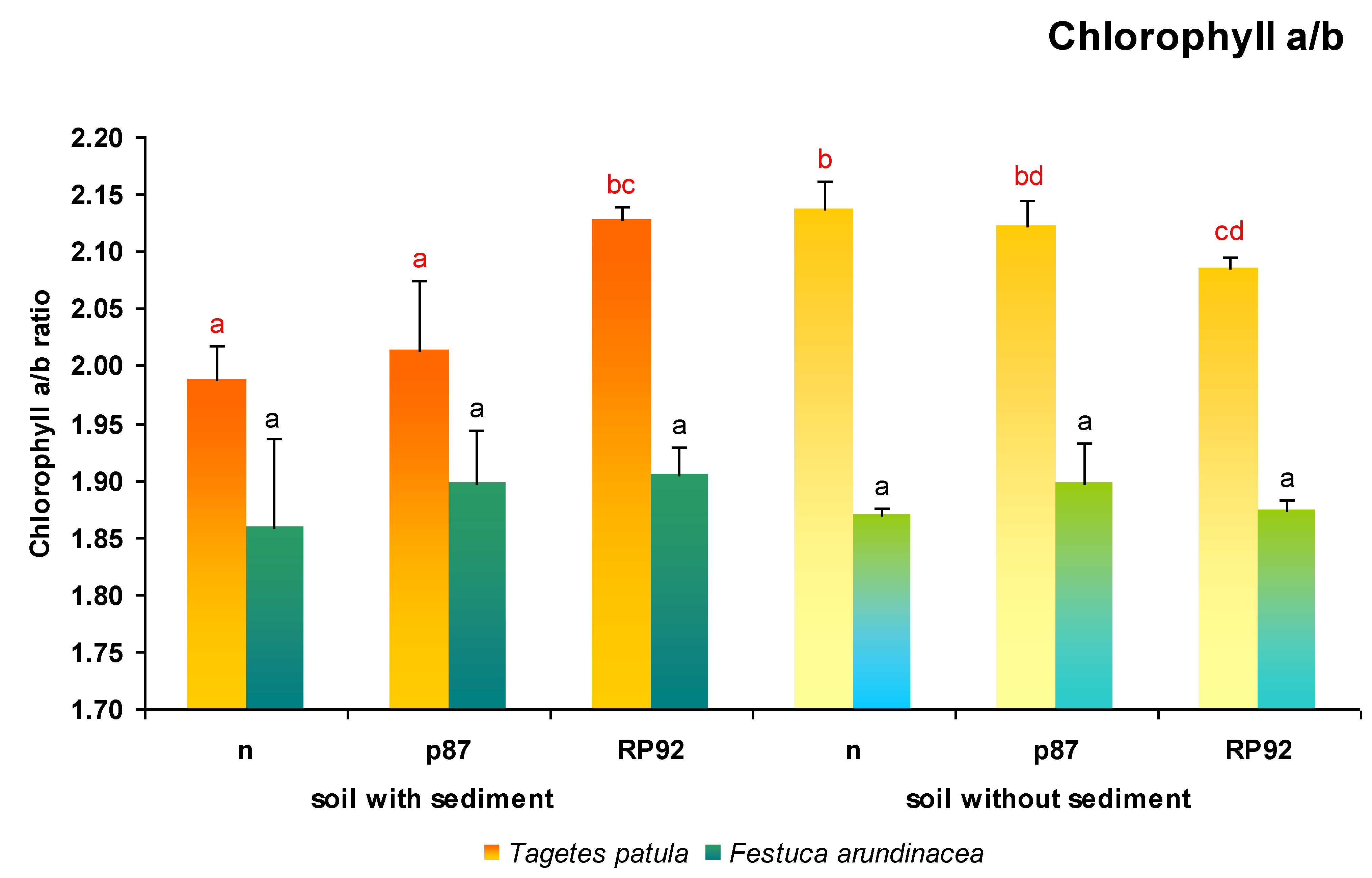
In
T. patula plants (
Figure 5), both the presence of sediments in soil (F = 31.00; df = 1, 18;
p = 0.000028) and the type of inoculation (F = 4.7; df = 2, 18;
p = 0.0224) influenced the chlorophyll a/b ratio (ANOVA II). For the soil treated with sediment, the non-inoculated plants and plants inoculated with p87 demonstrated significantly lower chlorophyll a/b ratio (from 7% to 3.5%) compared to those inoculated with RP92. The RP92 inoculated plants also demonstrated a lower chlorophyll a/b ratio than the non-inoculated plants on soil without sediments (2.5%) and those treated with PR92 on sediment-amended soil (2%).
In the case of F. arundinacea plants, no statistically significant differences were found between variants regarding the chlorophyll a/b ratio (ANOVA II).
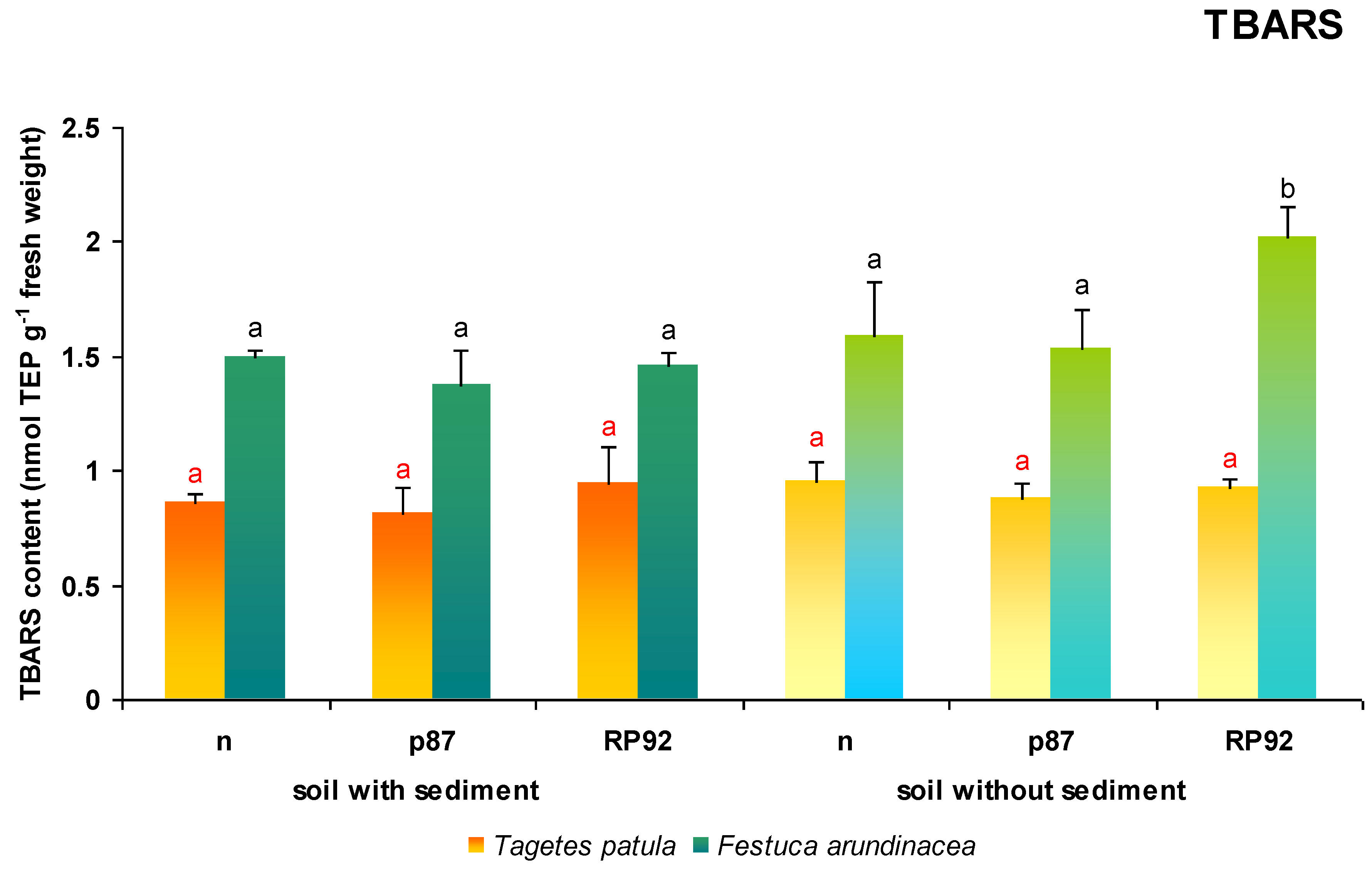
3.5. TBARS Content
Neither the presence of sediment nor the type of inoculation influenced the TBARS value in
T. patula leaves, an indicator of oxidative lipid damage (ANOVA II;
Figure 6).
In contrast, in the case of F. arundinacea, both the presence of sediment in soil (F = 20.647; df = 1, 18; p = 0.00025) and the type of inoculation influenced TBARS content in leaves (F = 7.863; df = 2, 18; p = 0.0035; ANOVA II). However, leaf tissue TBARS content only increased in the untreated soil inoculated with the RP92 strain, reaching between 127% and 146% of the value found in the other experimental variants (Duncan test).
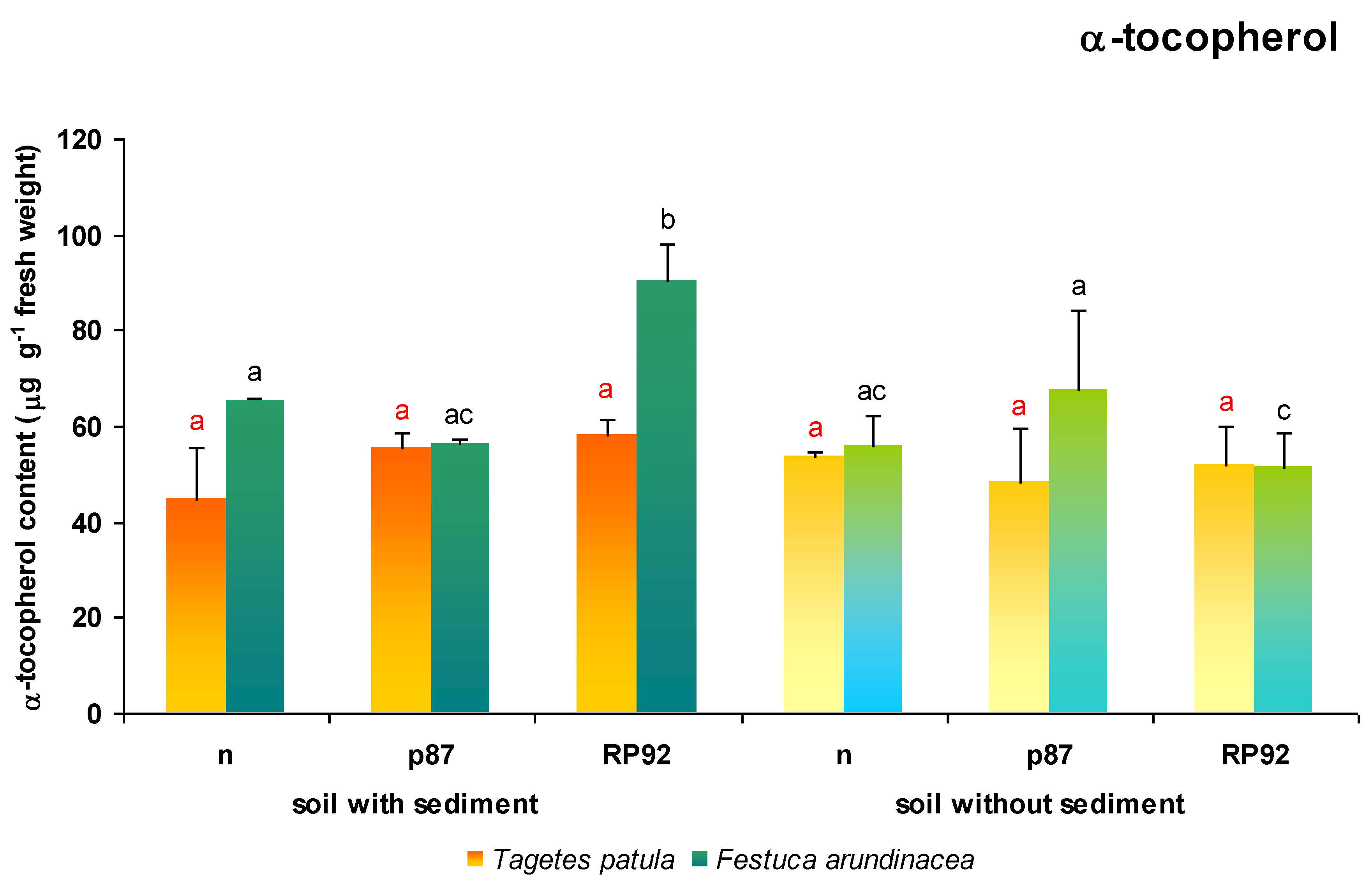
3.6. α-Tocopherol Content
No statistically significant differences in α-tocopherol content were observed between any pairs of experimental variants in
T. patula leaves (ANOVA II;
Figure 7).
The content of α-tocopherol, the main antioxidant of lipid fraction in F. arundinacea plants depended on both the presence of sediment (F = 13.039; df = 1, 18; p = 0.002) as well as the type of inoculation (F = 3.736; df = 2, 18; p = 0.044). However, the highest α-tocopherol content was demonstrated by the plants growing on the soil enriched with sediment and the inoculated strain RP92 (Duncan test), with its value ranging from 134% to 175% of those found in the other variants. In addition, in soil without sediments, α-tocopherol content was found to be 30% higher in soil treated with the p87 strain than with RP92. The latter also demonstrated 21% lower α-tocopherol content than plants growing on non-inoculated soil with sediments.
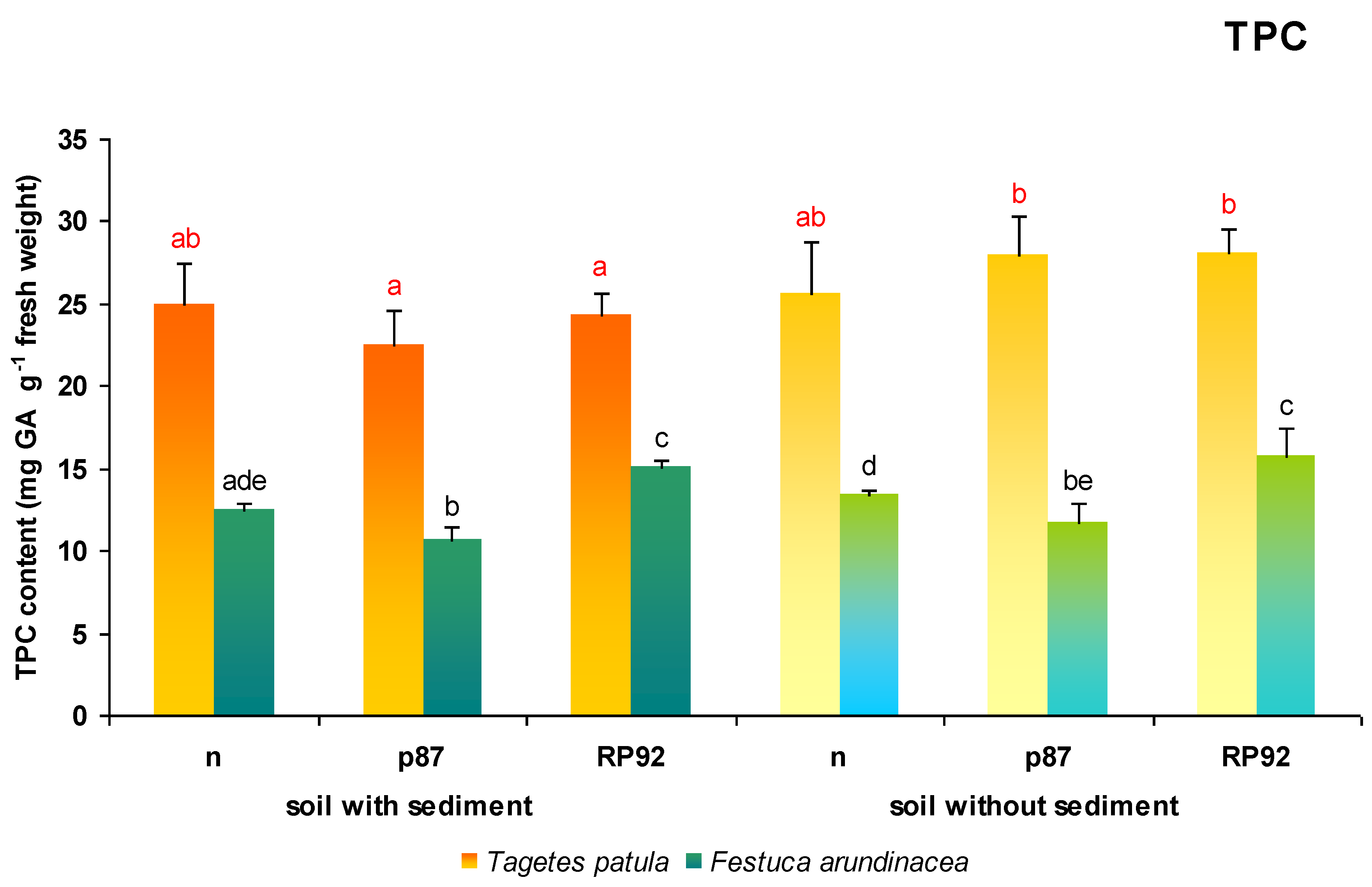
3.7. TPC Content
The presence of bottom sediments in the soil had a significant effect on the TPC content in
T. patula plants (ANOVA II; F = 13.41; df = 1, 18;
p = 0.00178;
Figure 8); however, the type of inoculation also influenced TPC content: On soil without sediments, plants inoculated with p87 demonstrated 124% higher TPC content and those inoculated with RP92 115% higher, than the p87 and RP92 plants growing in treated soil.
In F. arundinacea plants, both the presence of sediments in the soil (F = 4.747; df = 1, 18; p = 0.0423) and the type of inoculation had an effect on leaf TPC content (F = 39.556; df = 2, 18; p = 0.00000; ANOVA II). Differences were also found between the inoculated bacterial variants (Duncan post hoc test): Those treated with p87 in sediment-amended soil displayed 85% TPC compared to the plants growing on non-inoculated soil, while those treated with RP92 displayed 120% TPC. Similarly, in F. arundinacea plants growing on a substrate without sediments, inoculation with p87 strain decreased (88%), and inoculation with strain RP92 increased (118%) the TPC content in leaves compared to the plants growing on non-inoculated soil.
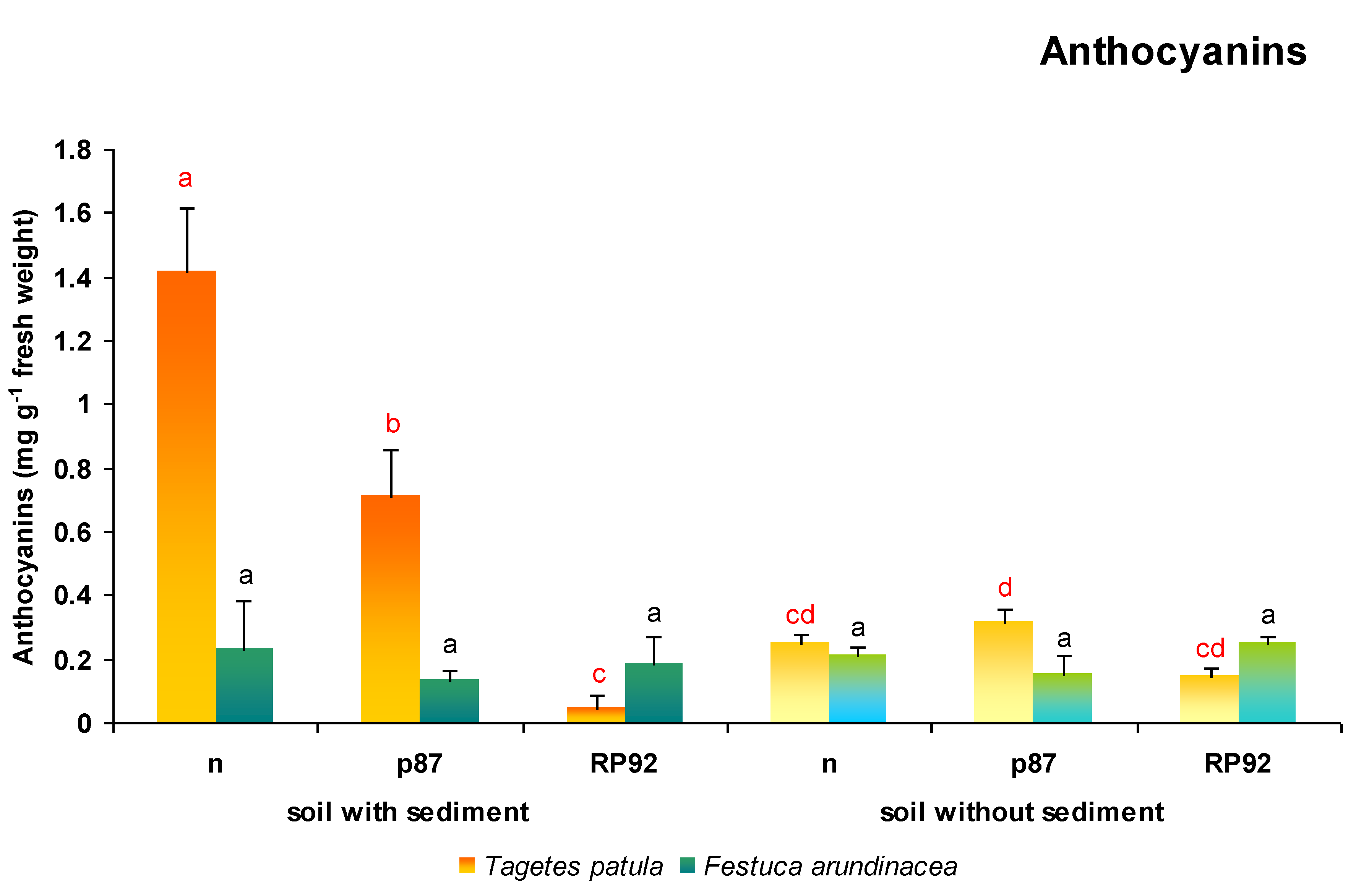
3.8. Anthocyanins Content
In
T. patula plants, both the presence of sediments in the soil (F = 66.248; df = 1, 18;
p = 0.0000) and the type of inoculation used had an effect on the content of anthocyanins in leaf tissues (F = 51.012; df = 2, 18;
p = 0.00000; ANOVA II;
Figure 9). On soil treated with sediment, non-inoculated variants demonstrated a 562% increase in anthocyanins content, and those inoculated with p87 displayed a 225% increase, compared to those grown on soil without sediments. In addition, anthocyanins content was lowest in plants growing on soil enriched with sediment and inoculated with RP92 strain: In this case, content was only 3.3% of that found in the variant with sediments, and 6.6% of the variant with sediment and p87 inoculation. Inoculation with bacterial strains did not appear to have any influence on the anthocyanins content of plants growing on soil without sediments.
In the case of F. arundinacea plants, neither the presence of sediment nor the type of inoculation used appeared to influence anthocyanins content.
4. Discussion
Based on changes in plant morphology, our findings suggest that the dicotyledonous
T. patula is more sensitive to the presence of substances included in urban sediments used to amend the soil in the present study than the monocotyledonous
F. arundinacea. In addition, inoculation of the soil with p87 and RP92 strains of bacteria reduces the toxic effects of this sediment on
T. patula. Previous studies show that morphological changes resulting from the same stress factor may vary significantly depending on the plant species. For example, one study found two species of Cucurbitaceae, zucchini and cucumber to display different reactions to sewage sludge treatment [
45]: Despite their relatedness, the leaves of the zucchini plants displayed chlorotic spots and bleaching, as well as necrosis and non-physiological shapes, while those of the cucumber plants showed no real damage. Nevertheless, despite such different reactions and different resistance to environmental stress factors, both species contributed to a similar degree to the reduction of substrate toxicity [
46]. The reduction in the severity of morphological changes observed after the application of bacterial strains p87 and RP92 to sediment-amended soil could be attributed to the fact that some PGPB strains may produce indole acetic acid, gibberellins, cytokinin, antioxidants and some unknown determinants, which may lead to an enhanced uptake of nutrients, thereby improving plant health under stress conditions and eliminating the effects of stress [
47].
In addition to morphological changes, inhibition of
T. patula growth and reduction of biomass yield after urban sediment use indicates their toxic effect on this plant species. Interestingly, the application of p87 and RP92 bacterial strains to soil enriched with sediments led to a 2.1 and 2.4-fold increase in biomass production, respectively, compared to plants growing on non-inoculated soils with sediments. In addition, the use of bacterial strains on untreated soil does not change the yield of biomass obtained: It is possible that if plants are not exposed to stress factors and, as a consequence, they develop properly with efficient biomass growth, the addition of bacterial strains to uncontaminated soil does not stimulate even greater biomass growth. This situation indicates that the presence of bacterial strains in the contaminated soil can trigger mechanisms in soil and/or in plants that allow the stress factor to interact directly with the plant or minimize its effects. The presence of PGPB in the soil facilitates plant growth by operating through two mechanisms: Indirectly by reducing plant pathogens and directly by facilitating nutrient uptake through phytohormone production, by enzymatic reduction of plant ethylene levels and/or by siderophore production [
47]. PGPB inoculation with
Pseudomonas mendocina in
Lactuca sativa plants affected by salt stress resulted in significantly greater shoot biomass than controls; the authors suggest that inoculation with selected PGPB could be an effective tool for alleviating salinity stress in salt sensitive plants [
48]. Moreover, when PGPB was used to seed bacterization, plants show enhanced root and shoot length, biomass and biochemical parameters such as chlorophyll, carotenoid and protein content [
49]. Otherwise excellent works on the remediation capabilities of
T. patula provide no information on the impact of using bacterial strains during remediation, or their significance for these plants [
31,
32]. Our present findings indicate that as in the case of plant morphology,
F. arundinacea did not show major changes in biomass yield following sediment treatment; however, the use of RP92 strain on soil without sediment had a promoting effect.
Differences in the biochemical parameters were also observed between T. patula and F. arundinacea, and these variations were related to the use of sediments and the presence of bacterial inoculums in the soil.
After the application of urban sediments,
T. patula plants responded with a decrease in total soluble protein, total chlorophyll and chlorophyll a/b ratio. Environmental factors that have an adverse effect on plant metabolism can lead to premature senescence, even before full plant maturity is achieved. One of the first symptoms of the aging process is a reduction in the concentration of total soluble protein in tissues; this has been observed even before the natural decline of photochemical processes in wheat leaves during the aging process [
50].
Total soluble protein and total chlorophyll content were also found to be key parameters among physiological and biochemical signs of senescence in a paper examining the influence of nitrogen deficit on aging in sugar beet [
51]. Their values were always lower in older leaves of sugar beet than in younger ones, and they decreased almost synchronously during leaf senescence. The authors attribute the observed decrease in total chlorophyll content to the destruction of the quickly renewing protein(s) of the chlorophyll-protein complex, resulting in immediate discoloration of the chloroplast.
The initial steps in the degradation of chlorophylls and chloroplast proteins can occur in intact organelles but a morphological analysis of senescing chloroplasts revealed the later breakdown of the thylakoid membrane system and degradation of thylakoid-bound proteins [
52]. Another indicator of the progressive senescence of green plant tissues is the presence of changes in chlorophyll a/b ratio. Nath et al., [
53] report a linear decrease in chlorophyll a/b ratio occurring simultaneously with a decrease in the total chlorophyll content in
Arabidopsis thaliana plants, which they attribute to different rates of degradation of chlorophyll a and chlorophyll b during senescence.
In various plants, the decrease of soluble protein commonly observed in the senescing leaves may also be related to proteolysis in the cell vacuoles [
54]. An increase in proteinase activity results in the degradation of proteins to amino acids; these are then transported through the phloem sap to the growing organs. This process of nitrogen remobilization plays an essential role at the whole-plant level during sequential senescence due to the fact that the mechanisms associated with senescence allow the redistribution of nutrients from old organs to sink organs for growth or storage [
55]. The correlation between nitrogen remobilization and senescence severity has been observed during the vegetative growth of several plants e.g.,
Arabidopsis [
56].
It is important to note that inoculation with the bacterial strain RP92 was found to increase the soluble protein content in the leaves of T. patula plants, both on those cultivated on sediment-amended soil, and those on unamended soil. In addition, in T. patula cultivated on sediment-amended soil, the use of RP92 almost doubles the chlorophyll content in the leaves and increases the chlorophyll a/b ratio. This may indicate that the strain has a protective effect on T. patula against the compounds in the applied sediments, which may act by preventing the plant from entering premature senescence. Of the two applied rhizobacterial strains, it seems that RP92 had a stronger protective effect than p87 on the analyzed parameters.
Among F. arundinacea plants, the use of urban sediments did not appear to cause significant changes in the content of soluble protein, but instead reduced the content of chlorophyll in green tissues. The use of bacterial strains generally had no effect on the soluble protein content of the tissues after sediment application, and in the case of plants growing on soil without sediments, only the simultaneous introduction of the p87 strain increased soluble protein content. Of the tested groups, the F. arundinacea growing on non-amended soil supplemented with p87 displayed the highest total chlorophyll content. In addition, RP92 supplementation also partially negated the unfavorable impact of sediments by increasing the content of the green assimilation dye. However, changes in total chlorophyll content were minor and had no effect on the chlorophyll a/b ratio.
One of the defenses employed by plants against environmental stress concerns the occurrence of secondary oxidative stress, characterized by an imbalance between the formation of reactive oxygen species (ROS) and their utilization by antioxidative systems. ROS such as superoxide anions (O
2−˙), hydroxyl radicals (
˙OH), peroxyl radicals (ROO
˙), singlet oxygen (
1O
2) and hydrogen peroxide (H
2O
2), are characterized by very high reactivity and cause oxidative damage to proteins, lipids, nucleic acids and other biologically important molecules [
57]. Such damage can lead to dysfunctions and irreversible degradation of molecules that are part of cellular structures and, consequently, inhibit the growth and development of plants. One group of plant secondary metabolites with non-enzymatic antioxidant properties is those based on phenolic compounds. Phenols exert their antioxidant properties through their ability to quench free radical reactions due to their potential to chelate metal ions through the properties of their nucleophilic aromatic ring [
58].
Phenolic compounds are also believed to inhibit membrane lipid peroxidation by “catching” alkoxyl radicals (RO
˙). Some phenols, particularly flavonoids, stabilize membranes by decreasing this fluidity due to the ability of phenolics to bind to some of the integral membrane proteins and phospholipids. Decreased fluidity in turn limits the diffusion of free radicals and reduces the peroxidation of membrane lipids. Some phenolic compounds (phenylpropanoids) are able to perform protective functions in plant cells by forming lignins and suberin, which mechanically reinforce the cell wall [
58,
59].
In the case of the
T. patula plants studied in this work, the addition of urban sediments to the soil did not affect the TPC content in the leaves. Differences associated with the addition of sediments to the soil were only visible in the variants in which bacterial strains were also applied. Interestingly, regarding plants treated with p87 and RP92, those cultivated on soil without sediments displayed greater TPC than those grown on soil with sediments. It is possible that in
T. patula plants, these small changes in the antioxidant component of TPC were sufficient to maintain the correct redox balance and prevent oxidative changes following sediment treatment or bacterial supplementation, as suggested by the lack of changes in the degree of lipid peroxidation measured as TBARS content. No significant changes in TBARS content have also been observed in previous studies conducted on cucumber plants growing on soil with the addition of urban sediments [
60].
Similarly, no TPC changes were associated with the application of urban sediments for F. arundinacea plants. Compared to the non-inoculated plant cultures, TPC decreased following p87 supplementation but increased following RP92 supplementation, both findings being irrespective of sediment addition. In this case, it may be surprising that the F. arundinacea plants growing on unamended soil and inoculated with RP92 demonstrated increased TPC content and elevated TBARS, which may indicate the induction of oxidative damage of lipids. It is possible that in this case the increase in the content of TPC did not compensate for the increase in oxidative reactions.
It cannot be ruled out that insufficiencies may exist in other elements of the antioxidant system, for example, changes in the α-tocopherol content of
F. arundinacea plants. α-tocopherol is the main antioxidant of the lipid fraction of the cell; it is synthesized exclusively in oxygenic photosynthetic organisms and constitutes more than 90% of the foliar tocopherols. This lipophilic antioxidant has two main antioxidant functions; firstly, the quenching of singlet oxygen generated mostly by triplet chlorophyll in photosystem II; and secondly, the scavenging of harmful radicals by donating electrons to various receivers, such as to a lipid peroxyl radical to prevent membrane lipid peroxidation reactions [
61]. In
F. arundinacea plants, α-tocopherol content was found to increase significantly in plants grown on amended soil inoculated with RP92; however, in the leaves of inoculated plants growing on unamended soil, the α-tocopherol content is similar that of plants on non-inoculated soils. This could be a reason for the oxidative damage of lipids formed in this experimental variant and hence, the observed increase in TBARS value.
Anthocyanins are blue, red or purple phenolic compounds that act as water-soluble pigments. They accumulate in the vacuoles of a wide range of cells and give color to flowers and fruits. In addition, when present in the leaves of plants exposed to environmental stressors, they may exert a protective influence by mitigating the effects of stress. At present, their main functions are considered to be antioxidants and sunscreens, mediators of ROS-induced signaling cascades, chelating agents for metals and metalloids, and delayers of leaf senescence, especially in plants growing under conditions of nutrient deficiency [
62,
63]. Foliar anthocyanins can protect chloroplasts from the adverse effect of excess light and have the potential to reduce both the incidence and severity of photooxidative damage. Moreover, anthocyanic leaves often display characteristics typical of leaves growing under shaded conditions, such as a lower chlorophyll a/b ratio compared to green leaves of the same plant species that do not contain high levels of anthocyanins [
64]. The protective function of anthocyanins can be of great importance when leaves grown under conditions of high light intensity simultaneously face other environmental stressors such as cold, drought, salinity or wounding, when the capacity of plants to usefully process radiant energy is severely constrained. The biosynthesis of anthocyanins in such conditions can represent one way to reduce the risk of photoinhibition [
65].
Among all studied biochemical parameters in
T. patula, anthocyanins production demonstrated the most pronounced changes after exposure to urban sediments and inoculation with bacterial strains. After the application of urban sediments, the content of anthocyanins was 5.6 times higher than in plants grown on soil without sediments. Considering the plant functions performed by anthocyanins, and considering that anthocyanins are also markers of environmental stress, such a large increase in their content may indicate the presence of a strong environmental stressor associated with the presence of substances from urban sediments in the soil. In this case, the increase in the content of anthocyanins could be associated with their antioxidative function. However, it cannot be ruled out that if urban sediments were used, the anthocyanins may have been more employed for their chelating properties, although this depends on the composition of the sediments and the potential risk of metal ions in the substrate, as well as the availability of these metals. Previous studies have found urban sediments to be moderately contaminated with zinc, and that other metals tested are present with acceptable limits [
35]. This may indicate that a strong reaction associated with a high accumulation of anthocyanins in
T. patula tissues may be associated with the presence of other substances acting as stress factors such as PAH, PCDD or PCDFs. It cannot be ruled out that the increased content of anthocyanins in
T. patula protects green plant tissues from photoinhibition, especially as, as already mentioned, the chlorophyll a/b ratio was found to decrease in plants exposed to urban sediments. Our findings also show that the bacterial strains p87 and RP92 demonstrated a positive effect in mitigating the effect of environmental stress, as indicated by an associated decrease in the content of anthocyanins, which serves as an indicator of the occurrence of stress factors. In addition, the RP92 strain seems to have a stronger effect in this respect.

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}