1. Introduction
Pseudo-nitzschia Peragallo is a chain-forming diatom genus that is widespread in all oceans of the world [
1]. A number of these diatom species of this genus are known producers of a neurotoxic amino acid, domoic acid (DA), which when accumulated via trophic transfer in the food-web can have deleterious (amnesic shellfish poisoning, ASP) and even fatal effects to several marine organisms and less frequently to humans [
2,
3,
4].
Pseudo-nitzschia is the only diatom genus among 27 dinoflagellates and 1 raphidophyte species that is listed as responsible for harmful algal events in upwelling regions [
5].
Pseudo-nitzschia is generally common in upwelling systems e.g., in California Current System [
6,
7,
8,
9,
10], in Iberian System [
11,
12,
13,
14], and Benguela upwelling system [
15,
16,
17].
Understanding the relationships between the abundance of these diatoms, DA, and the characteristics of the environment will provide valuable information on the variable(s) that might determine their growth, distribution, and DA production. Several authors [
18,
19,
20,
21,
22,
23,
24] have related these blooms with changes in nutrient availability, and therefore with upwelling events and eutrophication. On the west coast of USA, an area where
Pseudo-nitzschia is a common harmful bloom former, DA outbreaks that are caused by
P. australis have been associated with upwelling events and increased but in declining phase nutrients [
25,
26]. Other authors have also found a relationship between high
Pseudo-nitzschia spp. cell abundance and low-nutrient conditions [
27,
28,
29]. Several laboratory experiments [
30,
31,
32,
33,
34,
35] have addressed the silicic acid and/or phosphorus, iron, copper limitations as significant factors for the toxin production of
Pseudo-nitzschia species. The relationship between DA production and the nutritional status appears to be complex in natural populations, since they are often composed of several species [
36,
37], in fact some authors have not found obvious environmental triggers of DA production [
16]. However, it seems that the silicic acid supply [
7,
9,
20,
24,
28,
38,
39] plays an important role. More specifically for
P. australis, Anderson [
9] suggested that weight ratio values of Si(OH)
4:NO
3− below 2 are found to be linked to DA production from this species. Recently, Trick et al. [
40] showed that iron and copper limitations, and not macronutrient stress, are more essential for the DA production in natural populations in the Pacific North-West.
In addition to nutrient availability, salinity has also been reported as an important factor that affects
Pseudo-nitzschia abundance and/or DA production, mainly in estuarine systems [
23,
24,
26,
29,
41]. Regarding temperature, although
Pseudo-nitzschia species are observed in a wide range of temperatures, some authors have identified the optimal ranges for specific species [
23,
39]. Moreover, in a recent work, [
42] report that warming could lead to an increase of abundance and toxicity of
Pseudo-nitzschia spp. blooms in California coastal waters. Other environmental factors identified as affecting to
Pseudo-nitzschia spp. distribution and bloom dynamics are light intensity [
23,
39], photoperiod [
38], and factors controlling nutrient availability, such as river flow [
29], rainfall [
23,
38], or upwelling [
10,
13,
14].
Harmful algal events that were attributed to the genus
Pseudo-nitzschia are considered as a reoccurring phenomenon along the northern boundary of the Iberian–Canary current upwelling system. These blooms have become a focal point of numerous studies since the first time that they were recorded in the autumn of 1994 [
43] and they are occasionally associated with sporadic DA production, which occurs primarily towards the latter stages of bloom development [
26]. Although several species of
Pseudo-nitzschia have been recorded in the Galician coastal embayments referred to as
rias (NW Spain), only
P. australis has been shown to produce DA in the region [
44]. Toxigenic events due to blooms of
P. australis in this area have been recorded before DSP (Diarrheic Shellfish Poisoning) events [
45] in thermohaline stratified water masses.
There is considerable interest in the study of
Pseudo-nitzschia and the detection of DA in the Galician
rias due to the economic and social importance of the extensive mussel culture in this region and human health concerns [
46,
47]. As part of a routine monitoring programme, the Technological Institute for the Control of the Marine Environment of Galicia (INTECMAR) has organized sampling on a weekly basis in the Galician
rias, where, among other water parameters,
Pseudo-nitzschia spp. abundance and biotoxins in mussels and other molluscs are recorded. DA detection in this area has been mainly studied on shellfish extracts [
48,
49] and on
Pseudo-nitzschia cultures [
44,
50]. However, very limited information is available on the particulate DA concentrations (pDA) in seawater.
Chlorophyll
a (chl
a) concentration can be used as a phytoplankton biomass indicator, since it is common to almost taxonomic groups [
51]. Thus, chl
a maps that were derived from passive ocean colour sensors could be useful for identifying and monitoring high biomass blooms, although they do not provide information regarding species or toxicity [
52]. In Galicia, MERIS imagery has already proven to be an effective tool to map the spatial and temporal distribution of high biomass algae events during an upwelling cycle [
53]. MERIS provides images with a 300 m spatial resolution in nadir (Full Resolution –FR mode) and fifteen bands from visible to near infrared to support the monitoring of coastal zones [
54]. Although MERIS is not operative since 2012, Sentinel-3 OLCI provides continuity to the MERIS dataset since 2016, providing the same spatial resolution (300 m), but with more spectral bands (21 instead of 15) ranging from 400 nm to 1020 nm [
55].
The aim of this study is two-fold. Firstly, it attempts to develop a framework for the detection of potentially toxic Pseudo-nitzschia spp. blooms that are based on satellite data. Secondly, it aims to identify key variables affecting the formation and toxicity of Pseudo-nitzschia spp. blooms during different seasonally and meteorologically defined conditions, typical in upwelling systems.
3. Results
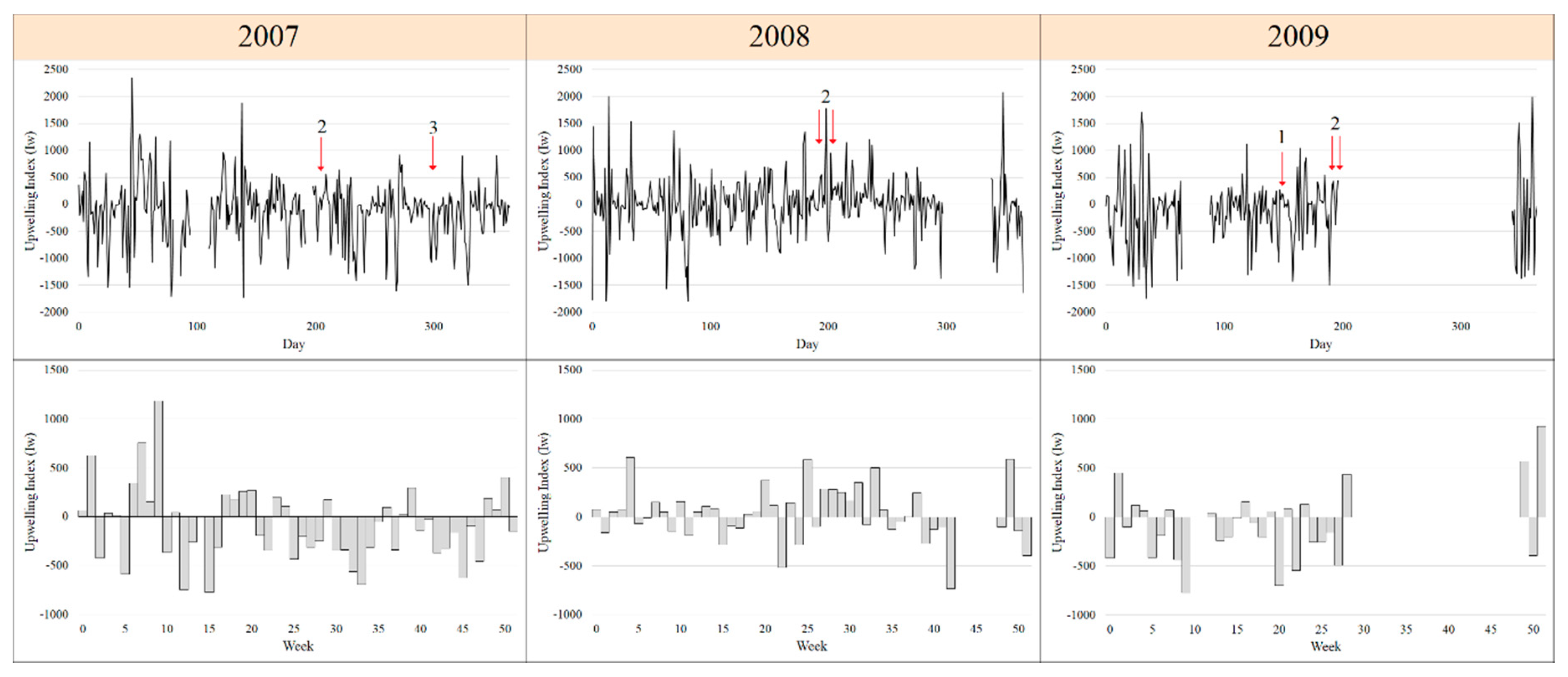
3.1. Environmental and Meteorological Conditions
Table 2 shows a list of the environmental parameters that were measured in this study and their mean values during the three periods. The surface concentration of nitrates ranged from below detection limit to 3.775 mg L
−1 and nitrites from 0.005 to 0.415 mg L
−1. Highest values for these two macronutrients were observed during the spring downwelling period. Orthophosphate at the sampling stations ranged from below detection limit to 0.850 mg L
−1, with high concentrations in the spring downwelling period. Silicic acid concentrations were generally low. The concentration levels of ammonium in surface waters in both studied
rias were generally low (below 0.023 mg L
−1), although elevated ammonium concentrations were observed in the Ria de Vigo during the summer period (22 July 2008). The POC surface concentration varied between 0.13 (autumn downwelling) and 0.75 mg L
−1 (summer). Higher levels of POC were observed in the summer period (22 July 2008). High concentrations of DOC were found in the autumn downwelling period in accordance with relatively high surface chl
a concentrations. The SPM concentrations varied from 1.17 to 3.74 mg L
−1 and the chl
a values ranged from 0.03 to 6.25 μg L
−1, with a maximum in the autumn downwelling period.
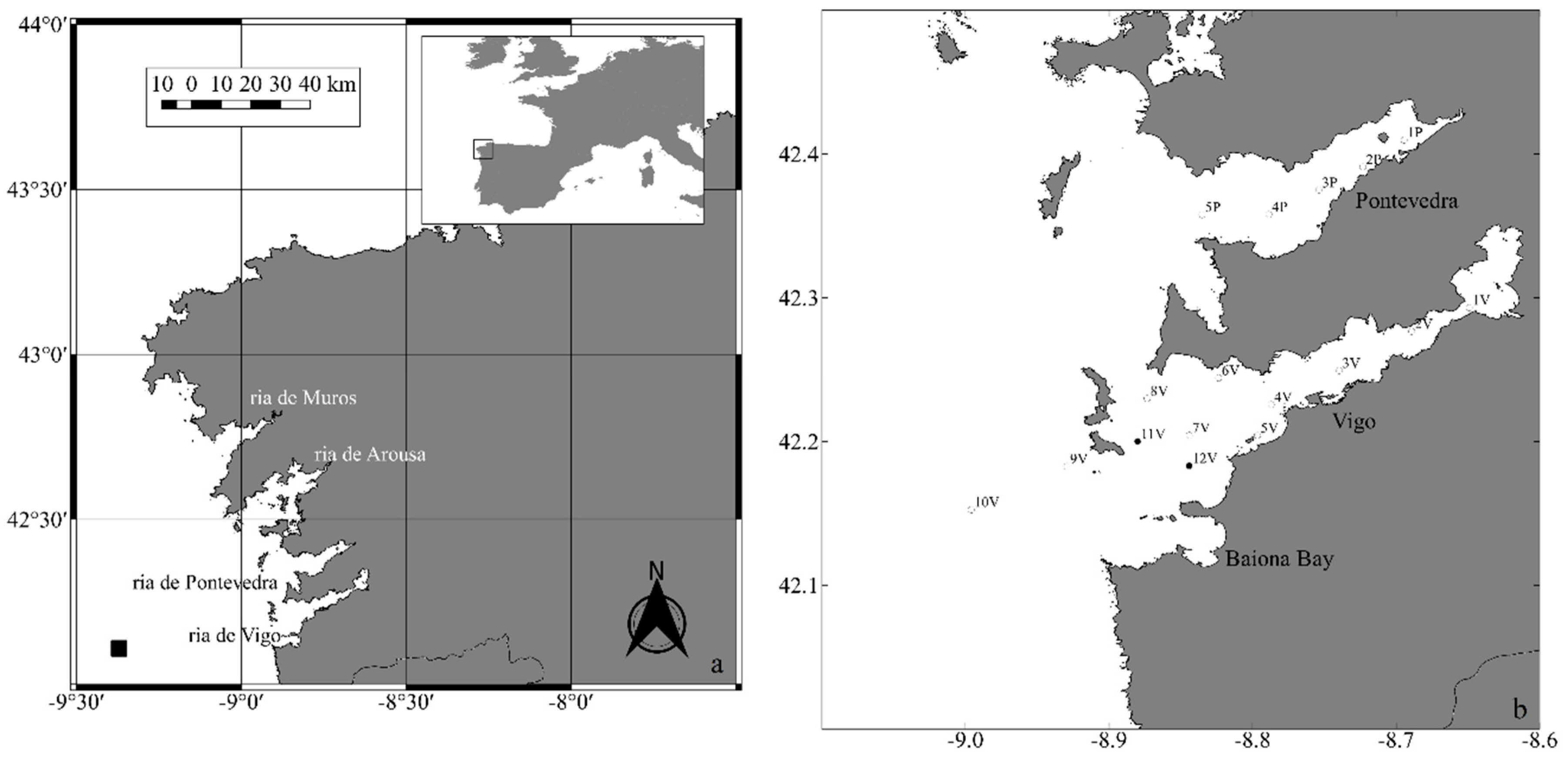
Samples from Ria de Pontevedra revealed higher concentrations of nitrate, nitrite, silicate, and orthophosphate, and the highest concentrations of the three first macronutrients were detected close to the main freshwater inputs in the ria (statistically significant at p < 0.05 for nitrite and orthophosphate). On the other hand, the ammonium and POC concentrations were generally higher in the Ria de Vigo with statistically significant differences only for ammonium. SPM concentrations showed, in general, decreasing values (in both rias) with distance from Station 1, which is located in the inner, narrow part of the rias and closer to the main freshwater inputs. In this part of the rias sediment resuspension and continental runoff is probably higher, resulting in high concentrations of SPM. ISM followed the same spatial and temporal distribution pattern as SPM. Secchi disk depths ranged from 2 m to 12 m in the Ria de Vigo and from 6 m to 11 m in the Ria de Pontevedra, generally less than half the water column depth.
3.2. Pseudo-nitzschia Composition, Abundance, and Particulate Domoic Acid
Scanning electron microscopy observations of the phytoplankton net samples revealed the dominance of
P. australis Frenguelli (
Figure 3).
P. pungens (
Figure 3) and
P. pseudodelicatissima were also found in the samples.
Pseudo-nitzschia was recorded during all of the surveys carried out in the study area and it was present in all of the samples.
Pseudo-nitzschia cells that belonged to the
P. delicatissima complex were detected in less than one-third of the samples, mainly in lower abundances than the
P. seriata complex and only when
Pseudo-nitzschia species belonging to
seriata complex were present.
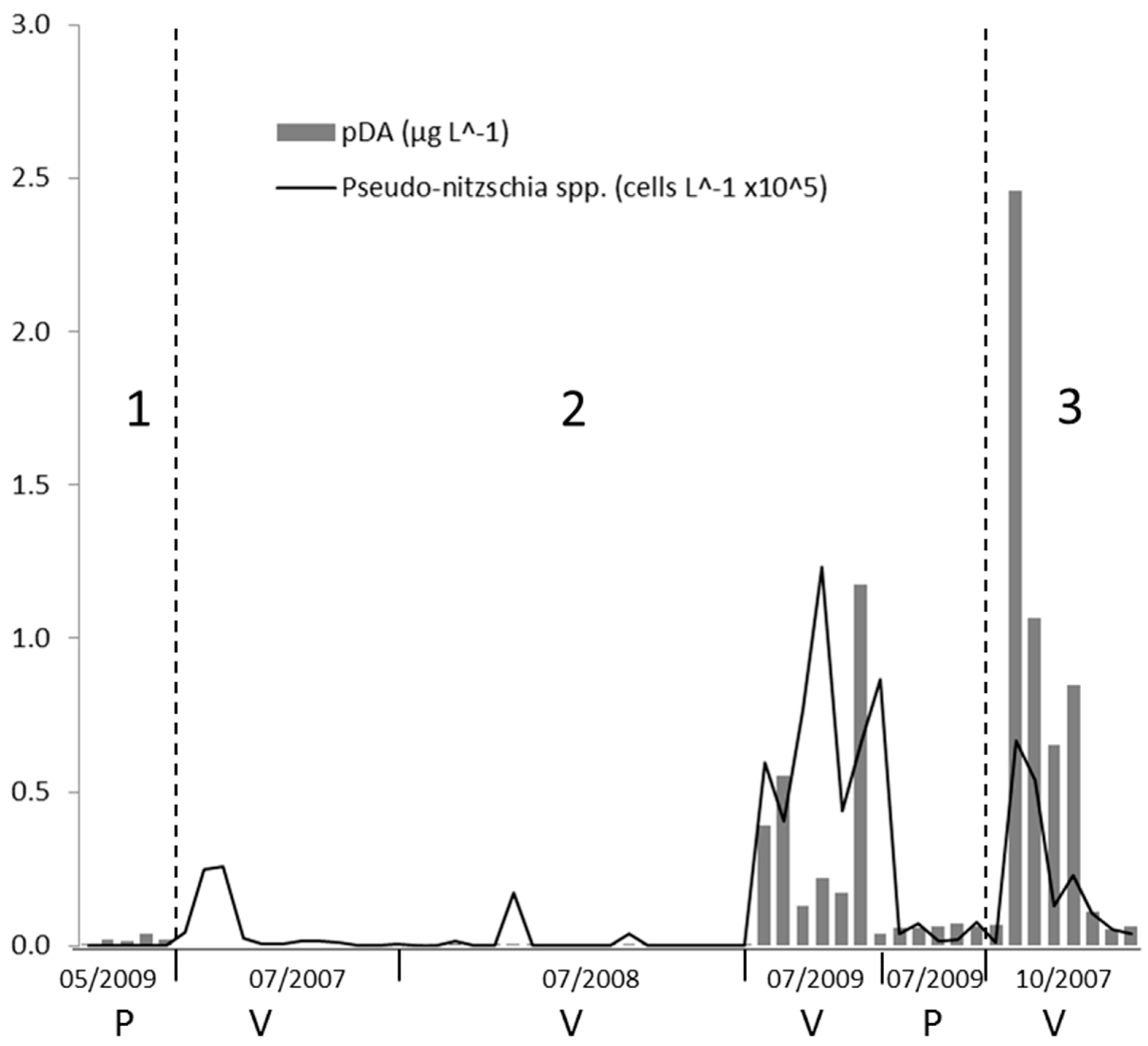
In 2008,
Pseudo-nitzschia spp. were recorded at only few stations and in general with low abundances while pDA was barely detectable in most of the samples (
Figure 4). Only
Pseudo-nitzschia species belonging to seriata complex were detected during these campaigns.
During the summer campaign in 2007, the maximum abundance of Pseudo-nitzschia spp. (> 0.44 104 cells L−1) in the Ria de Vigo was detected at the station 1, located at the innermost part of the ria.
In 2009,
Pseudo-nitzschia spp. abundances in the Ria de Pontevedra averaged 80 cells L
−1 on May 27 and 0.40 10
4 cells L
−1 on July 7. pDA values in all of the samples from the Ria de Pontevedra were above the detection limit, but relatively low (0.01–0.07 μg L
−1). The cellular DA levels varied from 0 to 54.31 pg DA cell
−1 (
Figure 4)
Pseudo-nitzschia spp. abundance almost linearly decreased from the warm surface waters (around 19 °C) at the inner part of the Ria de Vigo to the colder (17 °C) more saline waters outside the
ria. In order to investigate the dominant conditions during toxic blooms in more detail, the toxic
Pseudo-nitzschia events have also been individually studied in
Section 3.3.
3.3. Pseudo-nitzschia Toxic Events
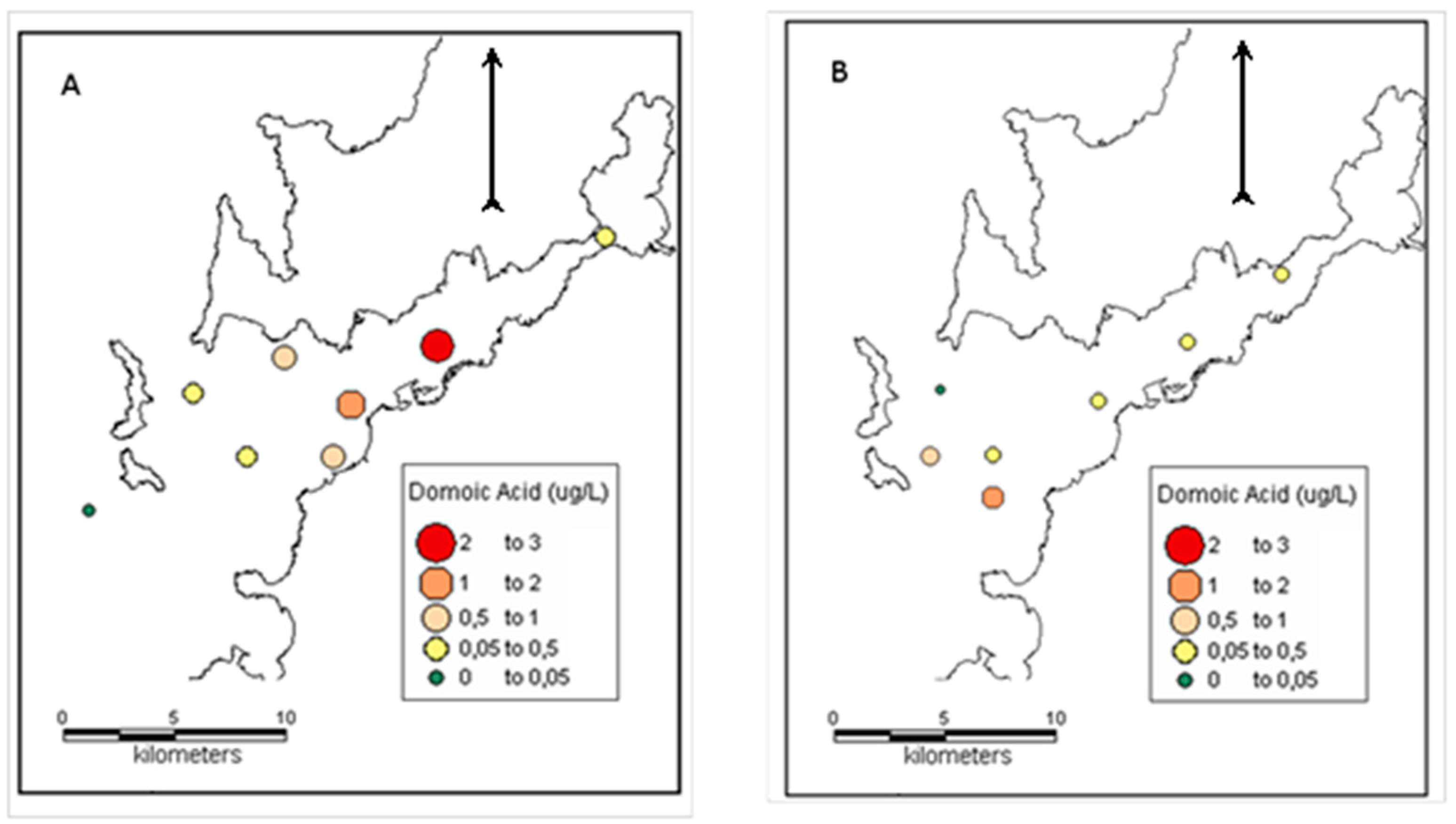
The first toxic event was recorded in the autumn downwelling period (19/10/2007) in the Ria de Vigo during a downwelling period. It was characterised by high abundances of
Pseudo-nitzschia spp. that belonged to the
seriata complex and DA concentrations up to 2.51 μg L
−1. Surface pDA was detectable at all of the sites with available samples. pDA concentrations were higher than 0.50 μg L
−1 throughout the middle part of the Ria de Vigo and they were still detectable, but in lower concentrations (close to 0.05 μg L
−1) outside and at the inner part of the
ria (
Figure 5). pDA in the Ria de Vigo showed a strong positive significant relationship with
Pseudo-nitzschia spp. in this dataset (Pearson’s
r = 0.92,
p = 0.001).
Pseudo-nitzschia spp. abundances and pDA concentrations were also highly correlated with Secchi disk depth (
r = −0.75), TSM (
r = −0.83), POC (
r = 0.87), chla (
r = −0.77), and the ratios of Si(OH)
4/PO
43− (
r = −0.72) and Si(OH)
4/N (
r = −0.71) (
Table 3). For the same survey, mean cellular DA was 31.7, ranging between 9.5 and 70.30 pg DA cell
−1. The highest cellular DA concentration was recorded at the site (V1) where the lowest
Pseudo-nitzschia spp. abundance (0.1 10
4 cells L
−1) was found.
On 14 July 2009 (summer period), a toxic event mostly due to
Pseudo-nitzschia seriata complex was monitored in the Ria de Vigo. During this event,
Pseudo-nitzschia spp. reached the highest abundance for the sampling period in the area (12 10
4 cells L
−1).
Pseudo-nitzschia abundance was high at all sites averaging 5.65·10
4 cells L
−1. pDA was detectable in all of the samples and ranged between 0.04 to 1.18 μg L
−1. The highest pDA concentrations were found in samples from sites close to the southern mouth of Ria de Vigo while the lowest concentration was recorded close to the northern mouth of the
ria (
Figure 5). The maximum pDA concentration for this sampling was measured in a sample containing 6.90·10
4 cells L
−1 of
Pseudo-nitzschia spp., while the minimum pDA concentration was found in a sample that contained the same magnitude of
Pseudo-nitzschia spp. (5.62·10
4 cells L
−1). Cellular DA concentrations averaged 14.20 pg DA cell
−1, showing a wide range of values (0.70–58.62 pg DA cell
−1). There was no significant correlation (Pearson’s test) between
Pseudo-nitzschia spp., DA concentration and cellular DA concentration for this sampling.
Pseudo-nitzschia spp. abundance was positively correlated to the Si(OH)
4/N ratio (
r = 0.75,
p = 0.05), while a strong relationship was revealed between the cellular DA concentration and N (
r = 0.96) and Si(OH)
4/N (
r = −0.92) (
Table 3).
3.4. Remote Sensing Detection
The FCM clustering method was applied in available MERIS FR images to build classification images defining the scope of the regionally specific algorithm for chl
a retrieval (NNRB). These images revealed large areas where Cluster#1 is dominant, and therefore NNRB could be applied to obtain reliable results of chl
a concentration.
Table 4 shows the percentage of pixels that belong to each cluster for each image over the Ria de Vigo and Ria de Pontevedra. Cluster#1 was dominant in both
rias, averaging almost 92% of the pixels in the six images, while Cluster#2 and Cluster#3 were assigned to an average of 7.9% and 0.3% of the pixels, respectively.
In most of the images, Cluster#1 is dominant, allowing for the chla mapping over large areas. Only on 14 July 2009, the relatively high percentage of pixels belonging to Cluster#2 in combination with the cloud cover averted a reliable continuous chla mapping of the rias.
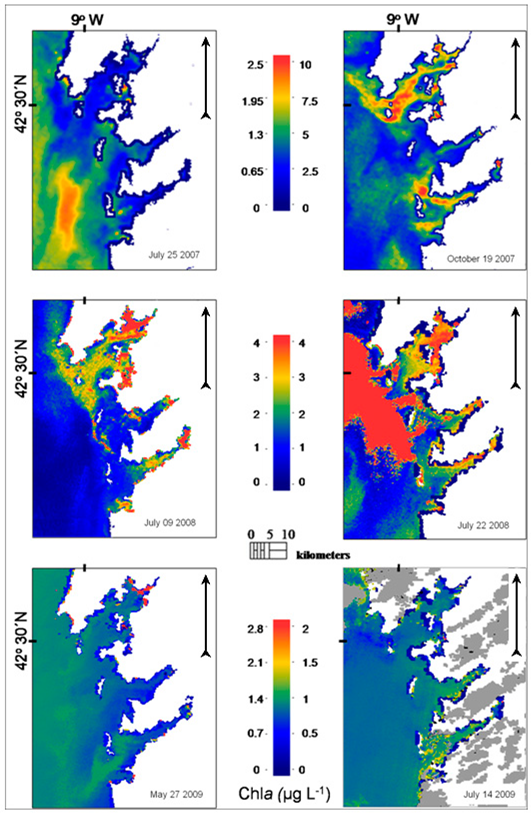
Spatial distribution of phytoplankton biomass in the study area was analysed from the available MERIS FR images while using the NNRB chl
a algorithm, showing heterogeneous results (
Figure 6). Elevated chl
a concentrations in the shallow innermost parts of the
rias observed in some images are most likely due to the high abundances of macroalgae on the remote sensing signal since in-situ measurements showed generally lower values.
On 25 July 2007, MERIS NNRB surface chla concentrations were relatively low (<1 μg L−1) in most parts of the Ria de Vigo and Ria de Pontevedra. An area of chla concentrations close to 2.5 μg L−1 was mapped in the area off the external coast of the rias. Generally, surface chla concentration was higher in offshore areas, where the lowest concentrations of Pseudo-nitzschia spp. were found.
Maximum MERIS NNRB chla concentrations were observed on 19 October 2007 coinciding with the first toxic event recorded during the sampling period and related to the presence of Pseudo-nitzschia spp. in the Ria de Vigo. In fact, the chla map shows high biomass “patches” clearly defined in this ria, i.e., zones of elevated chla concentrations (up to 10 μg L−1) at the middle part, just where the highest Pseudo-nitzschia spp. concentrations were measured, and also close to the northern and southern mouths. In the Ria de Pontevedra, the chla concentration was significantly lower in comparison with the concentrations observed in the Ria de Vigo and the Ria de Arousa. Areas of relatively high chla concentrations were noted in the adjacent area of the rias.
In the next MERIS image, which was acquired on 9 July 2008, a different chla distribution pattern is observed: the phytoplankton biomass is principally confined in the rias, while in the area off the rias the chla concentration remained at levels close to 0 μg L−1.
On 22 July 2008, high chla concentrations were mapped in the neighbouring shelf area. High concentrations were also extended to the outside part of Ria de Arousa and Ria de Pontevedra. The chla that was retrieved by NNRB in the Ria de Vigo showed wider variation than the concentrations measured in situ. During July 2008 Pseudo-nitzschia abundances remained at low levels.
On 27 May 2009, concentrations of chla lower than 1 μg L−1 were mainly noted in the three southern rias, while NNRB estimates were above this value off the rias. Low chla concentrations close to or lower than 1 μg L−1 were also observed both on the adjacent continental shelf and inside the rias on 14 July 2009.
The results of the multiple regression indicated that spectral reflectance in MERIS bands centred around 510 (Rrs_510), 560 (Rrs_560) and 620 (Rrs_620) nm explain 73% of the pDA variance (R2 = 0.73, F(2,32) = 32.26, p < 0.001). It was found that Rrs_510 significantly predicted pDA (β = 0.31, p < 0.001), as did Rrs_560 (β = −1.56, p < 0.01) and Rrs_620 (β = 3.24, p < 0.001).
3.5. Model Results of Pseudo-nitzschia spp. and pDA as a Function of Biotic and Abiotic Parameters
The results of generalised additive mixed models (GAMMs), which were used to model
Pseudo-nitzschia abundances in response to biotic and abiotic parameters, are summarised in
Table 5. The GAMMs results show a significant (
p < 0.005) linear relation of salinity and chl
a with
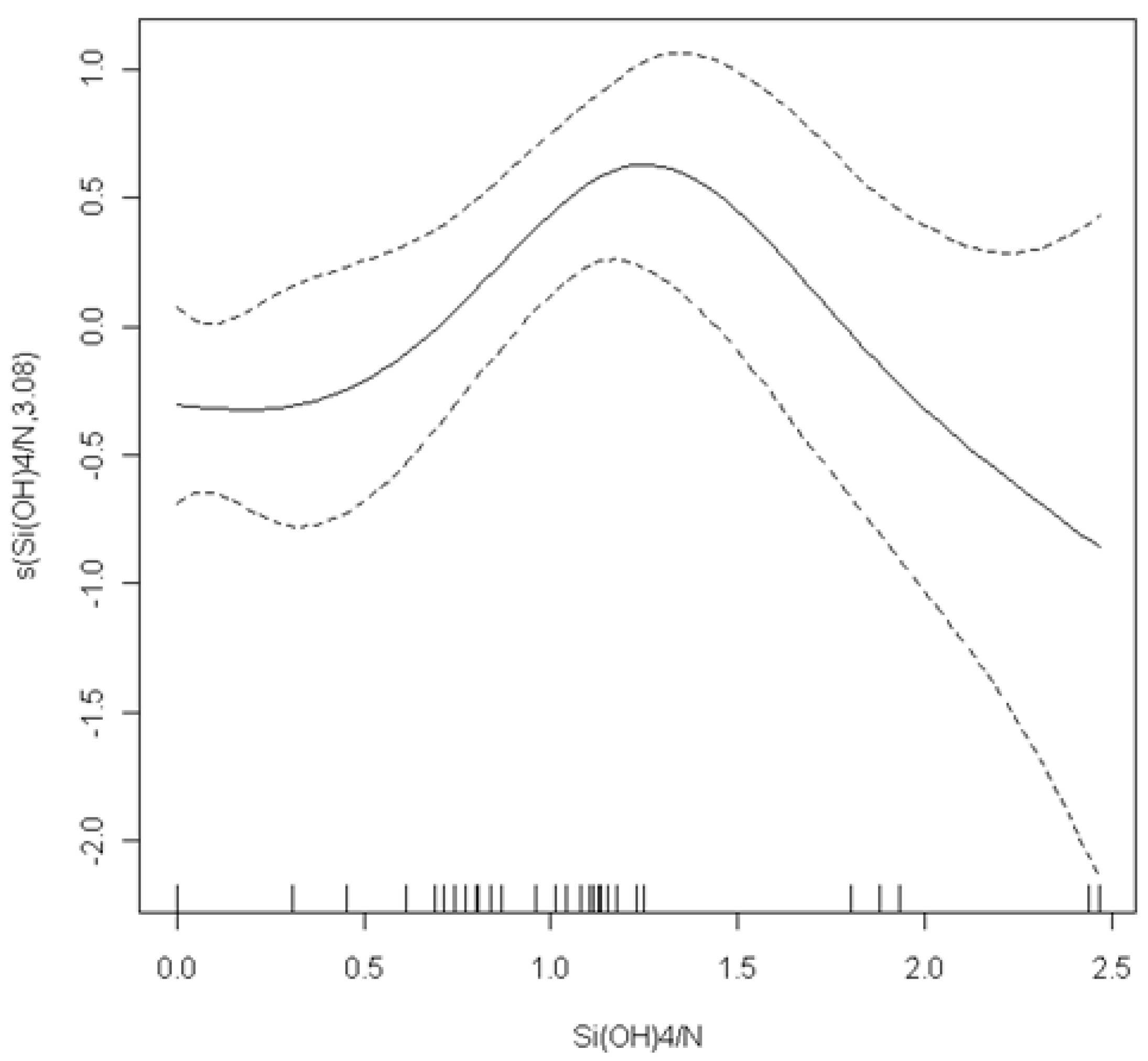
Pseudo-nitzschia abundance. The optimal model fitted the ratio Si(OH)
4/N in an additive way (
p = 0.02). The cross-validation estimated the degrees of freedom for this function to be 3.08. The smoothing curve that is shown in
Figure 7 suggests a peak of
Pseudo-nitzschia spp. abundance in moderate values of the Si(OH)
4/N ratio (close to 1.7) and an almost linear decrease for higher values. From all of the explanatory variables that were tested to model the pDA concentration, the final GLMM model included POC (
p = 0.02), NO
3− (
p = 0.03), PO
43− (
p = 0.004), and Si(OH)
4/N (
p = 0.02). Parameters estimates with SE and
t-values for the linear covariates are given in the
Table 6. Examination of the residuals did not indicate patterns.
5. Summary & Concluding Remarks
Domoic acid (DA) was measured in the seawater samples from the Galician
rias and revealed the presence of high levels in several cases. DA concentrations were not always related to
Pseudo-nitzschia abundances, showing that cell counts are not enough for assessing the toxicity of
Pseudo-nitzschia occurrence in the area. Despite the fact that no serious illnesses that were caused by ASP were reported in the area (Intecmar, Ministerio de Sanidad y Consumo) during the study period, DA levels, such as the ones detected here, could have the potential for significant impacts on the ecosystem and human health (e.g., by chronic exposure to moderate toxin levels, as it is mentioned by [
106] and [
107]). Furthermore, the DA content in some potential vectors that can be consumed by humans and other marine animals has not been studied in the Galician
rias.
In this study, mixed effect modelling was used to associate several environmental factors with the toxic Pseudo-nitzschia events for three different seasonal upwelling-downwelling conditions. The major aspects of phytoplankton succession in the area are relatively well known, in terms of major groups (diatoms, dinoflagellates, flagellates). Here, we intended to obtain to a finer degree of resolution, focusing in a particular group of diatoms. The results show that higher salinity values and lower, but not the lowest, Si(OH)4/N ratios favour higher Pseudo-nitzschia spp. abundances. High pDA values seem to be associated with relatively high PO43−, low NO3− concentrations, and low Si(OH)4/N. Knowledge of these relationships could help in evaluating the potential impacts of the blooms due to Pseudo-nitzschia on mariculture and wildlife, and improve our ability to predict toxic events as a function of environmental parameters.
The relationships that were found by the models only apply to the study area and the data that were collected during this period. Longer time-series from a wider area, samples from discrete depths, and micronutrients data could offer more detailed insight regarding the occurrence of toxic Pseudo-nitzschia spp. events. Nevertheless, this study documents the presence of toxigenic Pseudo-nitzschia and suggests that the monitoring of DA in the natural populations of Pseudo-nitzschia could be useful for identifying potential risks for DA contamination into higher trophic levels. Further research is also needed in the pathways of DA to highler trophic levels and examination of the relationships between Pseudo-nitzschia spp. blooms in the area and closures of mussel farms due to DA.
In this study, chl
a is retrieved from MERIS FR data while using a regional algorithm previously developed [
75]. A part of the chl
a data from the surveys described here has been used for the development of this regional neural network-based algorithm, which has been shown to permit the accurate mapping of chl
a of the
rias [
53]. In our MERIS imagery set, Cluster#1 was dominant in the three
rias and the adjacent area allowing for reliable chl
a mapping over the area. The autumn-time DA event was characterised by high chl
a concentrations. The highest
Pseudo-nitzschia spp. abundances (3 to 6 10
4 cells L
−1) were found in a relatively small area in the Ria de Vigo where elevated surface chl
a concentration (>6 mg m
−3) was mapped from the MERIS FR data. These small areas of high chl
a concentration accompanied with moderate abundances of toxic
Pseudo-nitzschia spp. can be easily missed by conventional monitoring techniques. The MERIS FR and MODIS (not shown here) images from the extended area of the Iberian Peninsula show very high chl
a concentrations, coupled with colder waters along the Portuguese and Galician coast indicating a general high biomass event.
Moreover, our results show “patchiness” in the spatial distribution of Pseudo-nitzschia in this coastal upwelling system, which suggests that the spatial and temporal resolution for the monitoring of these events should be increased. Ocean colour techniques can be helpful in tracking potentially harmful events that are caused by Pseudo-nitzschia and complement monitoring programs based on direct observations at fixed stations. Satellite data can be even more useful following the approach of this study with near real-time fine resolution imagery and regionally/cluster-specific algorithms for the retrieval of chla concentrations. According to the recorded in situ data, our approach to retrieve chla is an improvement on other previously used techniques, and made it possible to obtain reliable chla maps while using almost every image. The relationship that was established in this paper between the MERIS Rrs and pDA can be the basis of an effective Earth Observation (EO)-based HAB monitoring that will address the requirements of several key regional end-users.
Another finding is the relationship between MERIS FR data and pDA. In a previous study, Anderson et al. [
108] also developed regression models for
Pseudo-nitzschia spp. abundance, pDA, and cDa in the Santa Barbara channel integrating ocean colour (MODIS-Aqua and SeaWiFS) and sea surface temperature (AVHRR) data, finding relationships while using similar spectral bands (510 nm and 555 nm). Although our model is based on a limited dataset and it requires further research, it could be a good starting point for the development of new algorithms for the Rias Baixas area.
Although the Envisat satellite has stopped functioning in May 2012, the approach that was followed with MERIS by the authors could be adapted to other satellite-based ocean colour sensors. At present, we are involved in the European (EU H2020) funded project CoastObs [
109], which explores the potential use of the new Sentinel satellites to monitor coastal water environments. It includes, among its objectives, the development of validated map products that were derived from Sentinel 3 images that are expected to provide useful information for HABs detection and monitoring in Galicia, including not only chla concentration, but also species indicators for specific species, such as
Pseudo-nitzschia spp. or
Alexandrium minutum.

 and
and 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}