The Influence of Riparian Corridor Land Use on the Pesquería River’s Macroinvertebrate Community (N.E. Mexico)

 ,
,
Abstract
:1. Introduction
2. Methodology
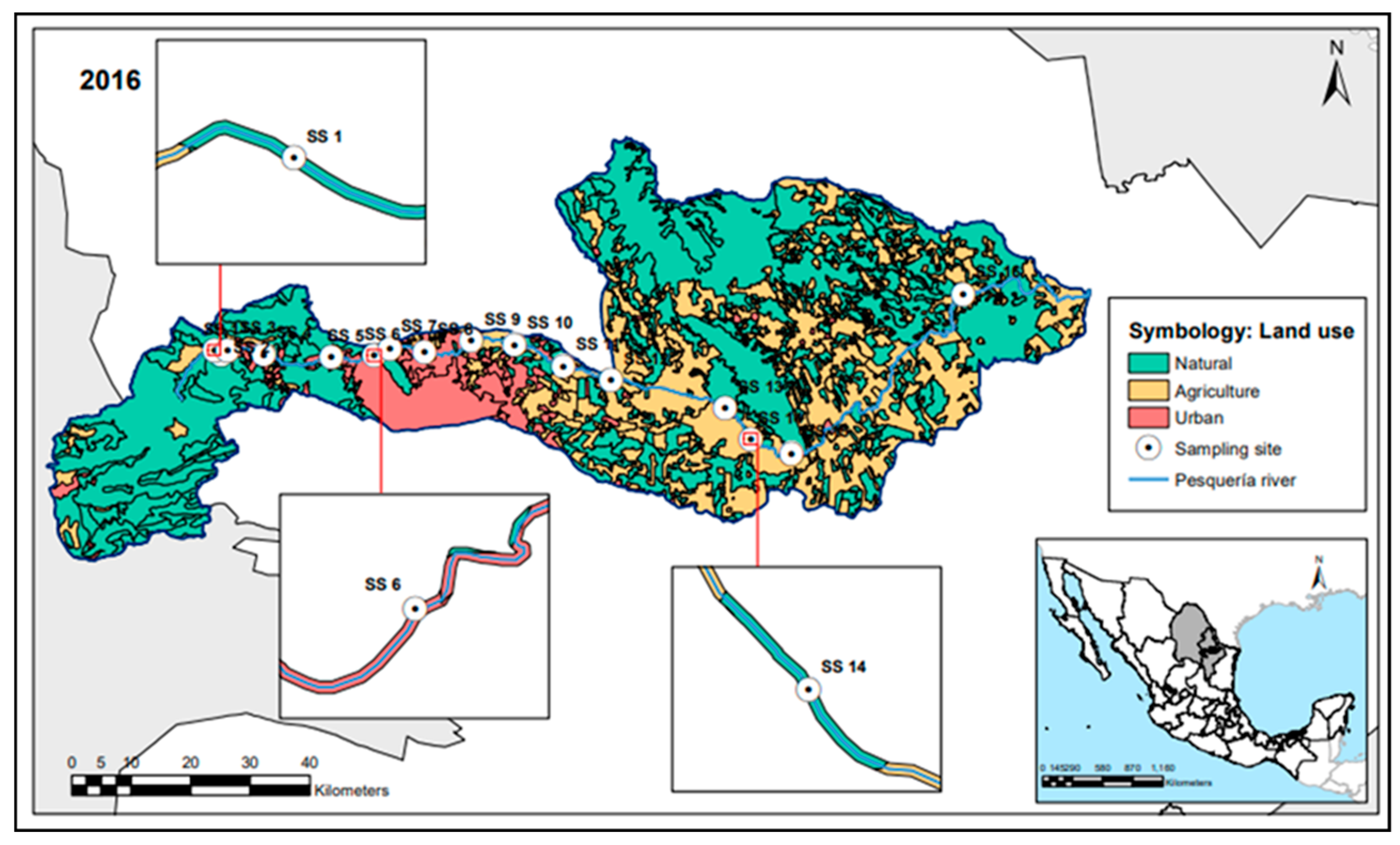
2.1. Study Area: The Pesquería River
2.2. Riparian Buffer Land Use and Sampling Site Characterization
2.3. Macroinvertebrate Sampling, Taxonomic Identification, and Biological Metrics
2.4. Data Analysis
3. Results
3.1. Land Use Characterization in the Pesquería River (PR)
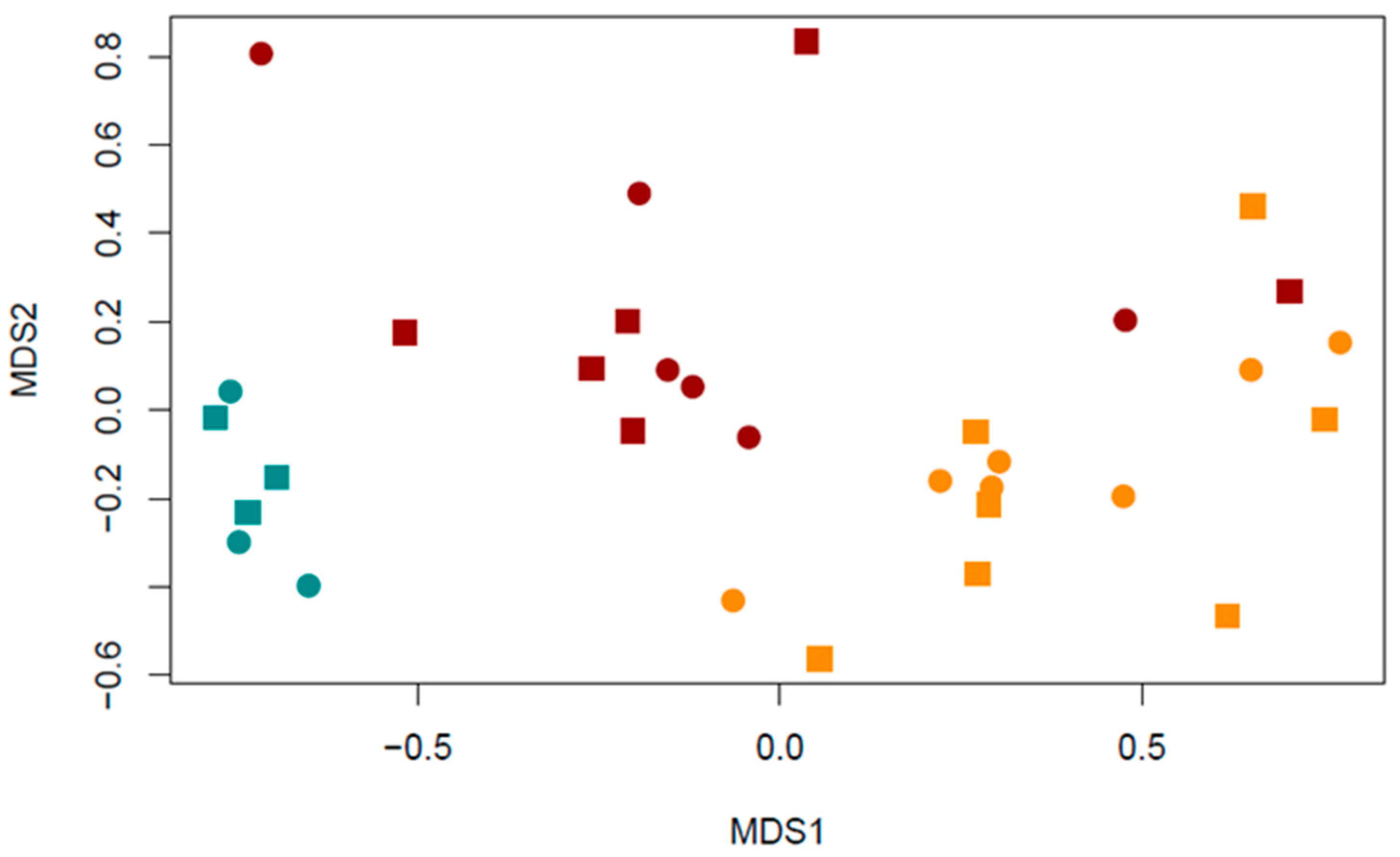
3.2. Aquatic Macroinvertebrate Community Structure and Land Use
3.3. Biological Metrics and Land Use
4. Discussion
5. Conclusions and Challenges
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sala, O.E.; Chapin, F.S.; Arme sto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Burdon, F.J.; Munz, N.A.; Reyes, M.; Focks, A.; Joss, A.; Räsänen, K.; Altermatt, F.; Eggen, R.I.L.; Stamm, C. Agriculture versus wastewater pollution as drivers of macroinvertebrate community structure in streams. Sci. Total Environ. 2019, 659, 1256–1265. [Google Scholar] [CrossRef] [PubMed]
- Price, E.L.; Sertić Perić, M.; Romero, G.Q.; Kratina, P. Land use alters trophic redundancy and resource flow through stream food webs. J. Anim. Ecol. 2019, 88, 677–689. [Google Scholar] [CrossRef]
- Media Release: Nature’s Dangerous Decline ‘Unprecedented’, Species Extinction Rates ‘Accelerating’|IPBES. Available online: https://www.ipbes.net/news/Media-Release-Global-Assessment#_ftnref1 (accessed on 11 May 2019).
- Krynak, E.M.; Yates, A.G. Benthic invertebrate taxonomic and trait associations with land use in an intensively managed watershed: Implications for indicator identification. Ecol. Indic. 2018, 93, 1050–1059. [Google Scholar] [CrossRef]
- McLaughlin, D.L.; Kaplan, D.A.; Cohen, M.J. A significant nexus: Geographically isolated wetlands influence landscape hydrology. Water Resour. Res. 2014, 50, 7153–7166. [Google Scholar] [CrossRef]
- Malmqvist, B.; Rundle, S. Threats to the running water ecosystems of the world. Environ. Conserv. 2002, 29, 134–153. [Google Scholar] [CrossRef]
- Walsh, C.J.; Roy, A.H.; Feminella, J.W.; Cottingham, P.D.; Groffman, P.M.; Morgan, R.P. The urban stream syndrome: Current knowledge and the search for a cure. J. N. Am. Benthol. Soc. 2005, 24, 706–723. [Google Scholar] [CrossRef]
- Stepenuck, K.F.; Crunkilton, R.L.; Wang, L. Impacts of urban landuse on macroinvertebrate communities in Southeastern Wisconsin streams1. Jawra J. Am. Water Resour. Assoc. 2002, 38, 1041–1051. [Google Scholar] [CrossRef]
- Death, R.G.; Baillie, B.; Fransen, P. Effect of Pinus radiata logging on stream invertebrate communities in Hawke’s Bay, New Zealand. N. Z. J. Mar. Freshw. Res. 2003, 37, 507–520. [Google Scholar] [CrossRef]
- Thompson, R.M.; Townsend, C.R. Land-use influences on New Zealand stream communities: Effects on species composition, functional organisation, and food-web structure. N. Z. J. Mar. Freshw. Res. 2004, 38, 595–608. [Google Scholar] [CrossRef]
- Lowrance, R.; Leonard, R.; Sheridan, J. Managing riparian ecosystems to control nonpoint pollution. J. Soil Water Conserv. 1985, 40, 87–91. [Google Scholar]
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef]
- Hollis, G.E. The effect of urbanization on floods of different recurrence interval. Water Resour. Res. 1975, 11, 431–435. [Google Scholar] [CrossRef]
- Pizzuto, J.E.; Hession, W.C.; McBride, M. Comparing gravel-bed rivers in paired urban and rural catchments of southeastern Pennsylvania. Geology 2000, 28, 79–82. [Google Scholar] [CrossRef]
- Gerhardt, A. Biomonitoring Polluted Water; Trans Tech Publications: Stafa-Zurich, Switzerland, 2000. [Google Scholar]
- Freshwater Biomonitoring and Benthic Macroinvertebrates; Rosenberg, D.M.; Resh, V.H. (Eds.) Springer: New York, NY, USA, 1993; ISBN 978-0-412-02251-7. [Google Scholar]
- Bonada, N.; Prat, N.; Resh, V.H.; Statzner, B. Developments in aquatic insect biomonitoring: A comparative analysis of recent approaches. Annu. Rev. Entomol. 2006, 51, 495–523. [Google Scholar] [CrossRef] [PubMed]
- Prat, N.; Ríos, B.; Acosta, R.; Rieradevall, M. Los Macroinvertebrados Como Indicadores de Calidad de las Aguas, 1st ed.; Domínguez, E., Fernández, H., Eds.; San Miguel de Tucumán Fundación Miguel Lillo: Tucumán, Argentina, 2009; ISBN 978-950-668-015-2. [Google Scholar]
- Villamarín, C.; Rieradevall, M.; Paul, M.J.; Barbour, M.T.; Prat, N. A tool to assess the ecological condition of tropical high Andean streams in Ecuador and Peru: The IMEERA index. Ecol. Indic. 2013, 29, 79–92. [Google Scholar] [CrossRef]
- Fierro, P.; Arismendi, I.; Hughes, R.M.; Valdovinos, C.; Jara-Flores, A. A benthic macroinvertebrate multimetric index for Chilean Mediterranean streams. Ecol. Indic. 2018, 91, 13–23. [Google Scholar] [CrossRef]
- Barbour, M.T.; Gerritsen, J.; Griffith, G.E.; Frydenborg, R.; McCarron, E.; White, J.S.; Bastian, M.L. A framework for biological criteria for Florida streams using benthic macroinvertebrates. J. N. Am. Benthol. Soc. 1996, 15, 185–211. [Google Scholar] [CrossRef]
- Plafkin, J.L.; Barbour, M.T.; Porter, K.D.; Gross, S.K.; Hughes, R.M. Rapid Bioassessment Protocols for Use in Streams and Rivers: Benthic Macroinvertebrate and Fish; Technincal Report No. EPA/444/4-89-001; Office of Water Regulations and Standards US EPA: Washington, DC, USA, 1989; pp. 1–34. [Google Scholar]
- Barbour, M.T.; Yoder, C.O. The multimetric approach to bioassessment, as used in the United States of America. Assessing the biological quality of fresh waters: RIVPACS and other techniques. In Proceedings of the International Workshop held in, Oxford, UK, 16–18 September 1997; pp. 281–292. [Google Scholar]
- Resh, V.H.; Norris, R.H.; Barbour, M.T. Design and implementation of rapid assessment approaches for water resource monitoring using benthic macroinvertebrates. Aust. J. Ecol. 1995, 20, 108–121. [Google Scholar] [CrossRef]
- Karr, J.R.; Chu, E.W. Biological Monitoring and Assessment: Using Multimetric Indexes Effectively; University of Washington: Wasahington, DC, USA, 1997. [Google Scholar]
- Heino, J. The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biol. Rev. 2013, 88, 166–178. [Google Scholar] [CrossRef]
- Delong, M.D.; Brusven, M.A. Macroinvertebrate community structure along the longitudinal gradient of an agriculturally impacted stream. Environ. Manag. 1998, 22, 445–457. [Google Scholar] [CrossRef]
- Rabení, C.F.; Doisy, K.E.; Zweig, L.D. Stream invertebrate community functional responses to deposited sediment. Aquat. Sci. 2005, 67, 395–402. [Google Scholar] [CrossRef]
- Pont, D.; Hughes, R.M.; Whittier, T.R.; Schmutz, S. A predictive index of biotic integrity model for aquatic-vertebrate assemblages of Western, U.S. Streams. Trans. Am. Fish. Soc. 2009, 138, 292–305. [Google Scholar] [CrossRef]
- Herman, M.R.; Nejadhashemi, A.P. A review of macroinvertebrate- and fish-based stream health indices. Ecohydrol. Hydrobiol. 2015, 15, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Mathuriau, C.; Silva, N.M.; Lyons, J.; Rivera, L.M.M. Fish and macroinvertebrates as freshwater ecosystem bioindicators in Mexico: Current state and perspectives. In Water Resources in Mexico: Scarcity, Degradation, Stress, Conflicts, Management, and Policy; Oswald Spring, Ú., Ed.; Hexagon Series on Human and Environmental Security and Peace; Springer: Berlin/Heidelberg, Germany, 2011; pp. 251–261. ISBN 978-3-642-05432-7. [Google Scholar]
- Arriaga Cabrera, L.; Aguilar Sierra, V.; Alcocer Durán, J. Aguas Continentales y Diversidad Biológica de México; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO): Mexico City, Mexico, 2000; ISBN 970-9000-15-2. [Google Scholar]
- Ruiz-Picos, R.A.; Kohlmann, B.; Sedeño-Díaz, J.E.; López-López, E. Assessing ecological impairments in Neotropical rivers of Mexico: Calibration and validation of the biomonitoring working party index. Int. J. Environ. Sci. Technol. 2017, 14, 1835–1852. [Google Scholar] [CrossRef]
- Ruiz-Picos, R.A.; Sedeño-Díaz, J.E.; López-López, E.; Ruiz-Picos, R.A.; Sedeño-Díaz, J.E.; López-López, E. Ensambles de macroinvertebrados acuáticos relacionados con diversos usos del suelo en los ríos Apatlaco y Chalma-Tembembe (cuenca del Río Balsas), México. Hidrobiológica 2016, 26, 443–458. [Google Scholar]
- Weigel, B.M.; Henne, L.J.; Martínez-Rivera, L.M. Macroinvertebrate-based index of biotic integrity for protection of streams in west-central Mexico. J. N. Am. Benthol. Soc. 2002, 21, 686–700. [Google Scholar] [CrossRef]
- Castro-López, D.; Guerra-Cobián, V.; Prat, N. The role of riparian vegetation in the evaluation of ecosystem health: The case of semiarid conditions in Northern Mexico: The riparian forest quality in arid rivers from the north of Mexico. River Res. Appl. 2019, 35, 48–59. [Google Scholar] [CrossRef]
- García, E. Modificaciones al Sistema de Clasificación Climática de Köppen; Universidad Nacional Autónoma de México: Mexico City, Mexico, 2004; ISBN 970-32-1010. [Google Scholar]
- Ferriño, A.L.F. Delimitación de zonas federales y áreas de amortiguamiento en ríos afectados por el crecimiento urbano como estrategia para prevenir inundaciones. Epistemus Ciencia Tecnología y Salud 2016, 19, 24–33. [Google Scholar]
- Anuario Estadístico y Geográfico de Nuevo León 2016; Anuario Estadístico y Geográfico de los Estados Unidos Mexicanos; Anuario Estadístico y Geográfico por Entidad Federativa; Síntesis Estadísticas Municipales; Instituto Nacional de Estadística y Geografía: Mexico City, Mexico, 2017; p. 614. ISBN 978-607-739-993-3.
- Torres Barajas, M.; Favela Lara, S.; Alanís Flores, G.; González Rojas, J.I.; Torres Barajas, M.; Favela Lara, S.; Alanís Flores, G.; González Rojas, J.I. Dinámica de uso de suelo en una región hidrológica prioritaria de la cuenca Río Bravo, Nuevo León. Revista Mexicana de Ciencias Forestales 2018, 9, 54–79. [Google Scholar] [CrossRef]
- Cañedo-Argüelles, M.; Kefford, B.J.; Piscart, C.; Prat, N.; Schäfer, R.B.; Schulz, C.-J. Salinisation of rivers: An urgent ecological issue. Environ. Pollut. 2013, 173, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Williams, W.D. Salinization of rivers and streams: An important environmental hazard. Ambio 1987, 15, 180–185. [Google Scholar]
- INEGI. Conjunto De Datos Vectoriales De Uso Del Suelo y Vegetación Escala 1:250,000, Serie VI (Capa Unión); Instituto Nacional de Estadística y Geografía (INEGI): Aguascalientes, Mexico, 2016. [Google Scholar]
- INEGI. Simulador de Flujos de Agua de Cuencas Hidrográficas; INEGI: Aguascalientes, Mexico, 2017. [Google Scholar]
- Munné, A.; Solá, C.; Pagés, J. Protocolo para la valoración de la calidad hidromorfológica de los ríos; Agencia Catalana de l Aigua: Barcelona, Spain, 2006; p. 82. [Google Scholar]
- Valera, C.A.; Pissarra, T.C.T.; Filho, M.V.M.; do Valle Júnior, R.F.; Oliveira, C.F.; Moura, J.P.; Sanches Fernandes, L.F.; Pacheco, F.A.L. The buffer capacity of riparian vegetation to control water quality in anthropogenic catchments from a legally protected area: A critical view over the Brazilian new forest code. Water 2019, 11, 549. [Google Scholar] [CrossRef]
- Broadmeadow, S.; Nisbet, T.R. The effects of riparian forest management on the freshwater environment: A literature review of best management practice. Hydrol. Earth Syst. Sci. Discuss. 2004, 8, 286–305. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Tomanova, S.; Tedesco, P.; Campero, M.; Van Damme, P.; Moya, N.; Oberdorff, T. Longitudinal and altitudinal changes of macroinvertebrate functional feeding groups in neotropical streams: A test of the river continuum concept. Fundam. Appl. Limnol. 2007, 170, 233–241. [Google Scholar] [CrossRef]
- Alanís-Rodríguez, E.; Jimenez-Pérez, J.; González-Rodríguez, H.; Canizales-Velázquez, P.A.; Mora-Olivo, A.; Mata Balderas, J.M.; Hernandez Salas, J. Composition, structure and diversity of shrublands in central Nuevo Leon, Mexico. Bot. Sci. 2015, 93. [Google Scholar] [CrossRef] [Green Version]
- Esri. ArcGIS 10.2 for Desktop 1999–2013 Esri Inc.; Environmental Systems Research Institute: Redlands, CA, USA, 2013. [Google Scholar]
- MAGRAMA. Protocolo de Muestreo y Laboratorio de Fauna Bentónica de Invertebrados en Ríos Vadeables; Secretaría General Técnica. Centro de Publicaciones: Madrid, Spain, 2013. [Google Scholar]
- Barbour, M.T.; Stribling, J.B.; Verdonschot, P.F.M. The multihabitat approach of USEPA’s rapid bioassessment protocols: Benthic macroinvertebrates. Limnética 2006, 25, 839–850. [Google Scholar]
- Domiguez, E.; Fernandez, H.R. Macroinvertebrados Bentónicos Sudamericanos; Fundacion Miguel Angel Lillo: Tucuman, Argentina, 2009; ISBN 978-950-668-015-2. [Google Scholar]
- Lanza-Espino, G.; de la Pulido, S.H. Organismos Indicadores de la Calidad del Agua y de la Contaminación (Bioindicadores); Plaza y Valdes: Madrid, Spain, 2000; ISBN 978-968-856-853-8. [Google Scholar]
- Merritt, R.W.; Cummins, K.W.; Daly, H.V.; Wiggins, G.B.; Berg, M.B.; Snider, R.J. An Introduction to Aquatic Insects of North America, 4th ed.; Kendall/Hunt Publishing Co, U.S.: Dubuque, IA, USA, 2008. [Google Scholar]
- Tachet, H. Invertébrés d’eau Douce: Systématique, Biologie, Écologie; CNRS Editions: Paris, France, 2010; ISBN 978-2-271-06945-0. [Google Scholar]
- Macedo, D.R.; Hughes, R.M.; Ferreira, W.R.; Firmiano, K.R.; Silva, D.R.O.; Ligeiro, R.; Kaufmann, P.R.; Callisto, M. Development of a benthic macroinvertebrate multimetric index (MMI) for neotropical savanna headwater streams. Ecol. Indic. 2016, 64, 132–141. [Google Scholar] [CrossRef]
- Edegbene, A.O.; Elakhame, L.A.; Arimoro, F.O.; Osimen, E.C.; Odume, O.N. Development of macroinvertebrate multimetric index for ecological evaluation of a river in North Central Nigeria. Environ. Monit. Assess 2019, 191, 274. [Google Scholar] [CrossRef]
- da Silveira de Oliveira, R.B.; Mugnai, R.; da Silva Pereira, P.D.; de Souza, N.F.; Baptista, D.F. A predictive multimetric index based on macroinvetebrates for Atlantic Forest wadeable streams assessment. Biota Neotropica 2019, 19. [Google Scholar] [CrossRef]
- Lu, K.; Wu, H.; Xue, Z.; Lu, X.; Batzer, D.P. Development of a multi-metric index based on aquatic invertebrates to assess floodplain wetland condition. Hydrobiologia 2019, 827, 141–153. [Google Scholar] [CrossRef]
- Tomanova, S.; Goitia, E.; Helešic, J. Trophic levels and functional feeding groups of macroinvertebrates in neotropical streams. Hydrobiologia 2006, 556, 251–264. [Google Scholar] [CrossRef]
- Ramírez, A.; Gutiérrez-Fonseca, P.E. Functional feeding groups of aquatic insect families in Latin America: A critical analysis and review of existing literature. Revista Biología Tropical 2014, 62, 155. [Google Scholar]
- Poff, N.L.; Olden, J.D.; Vieira, N.K.M.; Finn, D.S.; Simmons, M.P.; Kondratieff, B.C. Functional trait niches of North American lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. N. Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D. Vegan: Community Ecology Package. R package version 2.0-2. 2017. Available online: https://www.researchgate.net/publication/ 282247686 (accessed on 6 June 2019).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species:The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Thioulouse, J.; Dray, S.; Dufour, A.-B.; Siberchicot, A.; Jombart, T.; Pavoine, S. Multivariate Analysis of Ecological Data with Ade4; Springer: New York, NY, USA, 2018; ISBN 978-1-4939-8848-8. [Google Scholar]
- Roberts, D. Labdsv: Ordination and Multivariate Analysis for Ecology. R package version 1.7-0. 2015. Available online: https://github.com/cran/labdsv/commit/a56af17c747fd1480a0cac60818928776fc07edf (accessed on 5 May 2019).
- Feld, C.K.; Segurado, P.; Gutiérrez-Cánovas, C. Analysing the impact of multiple stressors in aquatic biomonitoring data: A cookbook with applications in R. Sci. Total Environ. 2016, 573, 1320–1339. [Google Scholar] [CrossRef]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Proceedings of the Second International Symposium on Information Theory, Akademiai Kaido, Budapest, Hungary, 2–8 September 1973; Akademiai Kaido: Budapest, Hungary, 1973. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009; ISBN 978-0-387-87458-6. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Gutiérrez-Cánovas, C.; Escribano-Ávila, G. Inferencia estadística a partir de varios modelos y su utilidad en ecología. Revista Ecosistemas 2019, 28, 118–120. [Google Scholar]
- Team R.C.R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Racine, J.S. RStudio: A platform-independent IDE for R and sweave. J. Appl. Econ. 2012, 27, 167–172. [Google Scholar] [CrossRef]
- Johnson, R.C.; Jin, H.-S.; Carreiro, M.M.; Jack, J.D. Macroinvertebrate community structure, secondary production and trophic-level dynamics in urban streams affected by non-point-source pollution. Freshw. Biol. 2013, 58, 843–857. [Google Scholar] [CrossRef]
- Hillebrand, H.; Bennett, D.M.; Cadotte, M.W. Consequences of dominance: A review of evenness effects on local and regional ecosystem processes. Ecology 2008, 89, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, J.C.; Kefford, B.J. Effects of Salinity on Stream Ecosystems: Improving Models for Macroinvertebrates; CSIRO: Canaberra, Australia, 2005. [Google Scholar]
- Velasco, J.; Millán, A.; Hernández, J.; Gutiérrez, C.; Abellán, P.; Sánchez, D.; Ruiz, M. Response of biotic communities to salinity changes in a Mediterranean hypersaline stream. Saline Syst. 2006, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Zinchenko, T.D.; Golovatyuk, L.V. Salinity tolerance of macroinvertebrates in stream waters (review). Arid Ecosyst. 2013, 3, 113–121. [Google Scholar] [CrossRef]
- Fierro, P.; Bertrán, C.; Tapia, J.; Hauenstein, E.; Peña-Cortés, F.; Vergara, C.; Cerna, C.; Vargas-Chacoff, L. Effects of local land-use on riparian vegetation, water quality, and the functional organization of macroinvertebrate assemblages. Sci. Total Environ. 2017, 609, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Acosta, R.; Ríos, B.; Rieradevall, M.; Prat, N. Propuesta de un protocolo de evaluación de la calidad ecológica de ríos andinos (CERA) y su aplicación a dos cuencas en Ecuador y Perú. Limnética 2009, 28, 35–64. [Google Scholar]
- Roldán, G. Revista de la Academia Colombiana de Ciencias Exactas Físicas y Naturales; Ministerio de Educación Nacional: Bogota, Columbia, 1999; pp. 375–387. [Google Scholar]
- Junqueira, V.M.; Campo, S.C.M. Adaptation of the BMWP for water quality evaluation to Rio das Velhas watershed (Minas Gerais, Brazil). Acta Limnologica Brasiliense 1998, 10, 125–135. [Google Scholar]
- Santiago-Fragoso, S.; Sandoval-Manrique, J.C. Coleópteros acuáticos y su relación con la dinámica fisicoquímica del Río Cuautla (Tramo Tetelcingo-Anenecuilco), Morelos, México. Hidrobiológica 2001, 11, 19–29. [Google Scholar]
- Twisk, W.; Noordervliet, M.A.W.; ter Keurs, W.J. Effects of ditch management on caddisfly, dragonfly and amphibian larvae in intensively farmed peat areas. Aquat. Ecol. 2000, 34, 397–411. [Google Scholar] [CrossRef]
- Koperski, P. Testing the suitability of leeches (Hirudinea, Clitellata) for biological assessment of lowland streams. Pol. J. Ecol. 2005, 53, 65–80. [Google Scholar]
- Burcher, C.L.; Benfield, E.F. Physical and biological responses of streams to suburbanization of historically agricultural watersheds. J. N. Am. Benthol. Soc. 2006, 25, 356–369. [Google Scholar] [CrossRef]
- Roy, A.H.; Rosemond, A.D.; Paul, M.J.; Leigh, D.S.; Wallace, J.B. Stream macroinvertebrate response to catchment urbanisation (Georgia, U.S.A.). Freshw. Biol. 2003, 48, 329–346. [Google Scholar]
- Kefford, B.J.; Marchant, R.; Schäfer, R.B.; Metzeling, L.; Dunlop, J.E.; Choy, S.C.; Goonan, P. The definition of species richness used by species sensitivity distributions approximates observed effects of salinity on stream macroinvertebrates. Environ. Pollut. 2011, 159, 302–310. [Google Scholar] [CrossRef]
- Short, T.M.; Black, J.A.; Birge, W.J. Ecology of a saline stream: Community responses to spatial gradients of environmental conditions. Hydrobiologia 1991, 226, 167–178. [Google Scholar] [CrossRef]
- Gallardo-Mayenco, A. Freshwater macroinvertebrate distribution in two basins with different salinity gradients (Guadalete and Guadaira river basins, south-western Spain). Int. J. Salt Lake Res. 1994, 3, 75–91. [Google Scholar] [CrossRef]
- Kay, W.R.; Halse, S.A.; Scanlon, M.D.; Smith, M.J. Distribution and environmental tolerances of aquatic macroinvertebrate families in the agricultural zone of southwestern Australia. J. N. Am. Benthol. Soc. 2001, 20, 182–199. [Google Scholar] [CrossRef]
- Naranjo López, C.; González Lazo, D.D.; Garcés González, G.; Brandimarte, A.L.; Muñoz Riveaux, S.; Musle Cordero, Y. Una metodología rápida y de fácil aplicación para la evaluación de la calidad del agua utilizando el índice BMWP-Cub para ríos cubanos. Tecnura 2005, 9, 65–76. [Google Scholar]
- Kholmogorova, N.V. Transformation Macrozoobenthos in the Small Rivers of Udmurtia Affected by Oil Mining, Extended Abstract of Cand. Sci. (Biol.). Ph.D. Dissertation, Kazan Federal University, Kazan, Russia, 2009. [Google Scholar]
- Piscart, C.; Moreteau, J.-C.; Beisel, J.-N. Biodiversity and structure of macroinvertebrate communities Along a small permanent salinity gradient (Meurthe River, France). Hydrobiologia 2005, 551, 227–236. [Google Scholar] [CrossRef]
- Figueroa, R.; Palma, A.; Ruiz, V.; Niell, X. Análisis comparativo de índices bióticos utilizados en la evaluación de la calidad de las aguas en un río mediterráneo de Chile: Río Chillán, VIII región. Revista Chilena de Historia Natural 2007, 80, 225–242. [Google Scholar] [CrossRef]
- Stewart, K.W. Chapter 205—Plecoptera: Stoneflies. In Encyclopedia of Insects, 2nd ed.; Academic Press: San Diego, CA, USA, 2009. [Google Scholar]
- Sánchez-Fernández, D.; Abellán, P.; Mellado, A.; Velasco, J.; Millán, A. Are water beetles good indicators of biodiversity in mediterranean aquatic ecosystems The case of the segura river basin (SE Spain). Biodivers. Conserv. 2006, 15, 4507–4520. [Google Scholar] [CrossRef]
- Moreno, J.L.; Suárez, M.L.; Vidal-Abarca, M.R. Valor ecológico de las ramblas como sistemas acuáticos singulares. In ‘Tomo extraordinario, 125 Aniversario de la Real Sociedad Española de Historia Natural’; Real Sociedad Española de Historia Natural: Madrid, Spain, 1996; pp. 411–414. [Google Scholar]
- Abellán, P.; Sánchez-Fernández, D.; Velasco, J.; Millán, A. Assessing conservation priorities for insects: Status of water beetles in southeast Spain. Biol. Conserv. 2005, 121, 79–90. [Google Scholar] [CrossRef]
- Kefford, B.J.; Buchwalter, D.; Cañedo-Argüelles, M.; Davis, J.; Duncan Richard, P.; Hoffmann, A.; Thompson, R. Salinized rivers: Degraded systems or new habitats for salt-tolerant faunas. Biol. Lett. 2016, 12, 20151072. [Google Scholar] [CrossRef] [PubMed]
- Kefford, B.J.; Papas, P.J.; Nugegoda, D. Relative salinity tolerance of macroinvertebrates from the Barwon River, Victoria, Australia. Mar. Freshw. Res. 2003, 54, 755–765. [Google Scholar] [CrossRef] [Green Version]
- MINAE. Reglamento Para la Evaluación y Clasificación de la Calidad de Cuerpos de Agua Superficiales; Technical Report No. 33903; La Gaceta: San José, Costa Rica, 2007; pp. 1–21. [Google Scholar]
- Davis, J.R. The creeping water bugs (Hemiptera: Naucoridae) of Texas. Southwest. Nat. 1996, 41, 1–26. [Google Scholar]
- Alba Tercedor, J.; Jáimez-Cuéllar, P.; Álvarez, M.; Avilés, J.; Bonada i Caparrós, N.; Casas, J.; Mellado, A.; Ortega, M.; Pardo, I.; Prat i Fornells, N.; et al. Caracterización del estado ecológico de ríos mediterráneos ibéricos mediante el índice IBMWP (antes BMWP’). Limnetica 2002, 21, 175–185. [Google Scholar]
- Ríos-Touma, B.; Acosta, R.; Prat, N. The andean biotic index (ABI): Revised tolerance to pollution values for macroinvertebrate families and index performance evaluation. Rev. De Biol. Trop. 2014, 62, 249–273. [Google Scholar] [CrossRef]
- Alvarez, L.F.; Roldan, G. Estudio del orden Hemiptera (Heteroptera) en el departamento de Antioquia en diferentes pisos altitudinales. Actualidades Biológicas 1983, 12, 31–46. [Google Scholar]
- Paul, M.J.; Meyer, J.L. Streams in the Urban Landscape. Annu. Rev. Ecol. Syst. 2001, 32, 333–365. [Google Scholar] [CrossRef]
- Freeman, P.L.; Schorr, M.S. Influence of watershed urbanization on fine sediment and macroinvertebrate assemblage characteristics in Tennessee Ridge and valley streams. J. Freshw. Ecol. 2004, 19, 353–362. [Google Scholar] [CrossRef]
- Dyer, S.D.; Wang, X. A comparison of stream biological responses to discharge from wastewater treatment plants in high and low population density areas. Environ. Toxicol. Chem. 2002, 21, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Serrano Balderas, E.C.; Grac, C.; Berti-Equille, L.; Armienta, H.M.A. Potential application of macroinvertebrates indices in bioassessment of Mexican streams. Ecol. Indic. 2016, 61, 558–567. [Google Scholar] [CrossRef]
- Cummins, K.W. Structure and function of stream ecosystems. BioScience 1974, 24, 631–641. [Google Scholar] [CrossRef]
- Kerans, B.L.; Karr, J.R. A benthic index of biotic integrity (B-IBI) for rivers of the Tennessee valley. Ecol. Appl. 1994, 4, 768–785. [Google Scholar] [CrossRef]
- Wallace, J.B.; Webster, J.R. The role of macroinvertebrates in stream ecosystem function. Annu. Rev. Entomol. 1996, 41, 115–139. [Google Scholar] [CrossRef] [PubMed]
- Miserendino, M.L.; Masi, C.I. The effects of land use on environmental features and functional organization of macroinvertebrate communities in Patagonian low order streams. Ecol. Indic. 2010, 10, 311–319. [Google Scholar] [CrossRef]
- Miserendino, M.L.; Pizzolon, L.A. Interactive effects of basin features and land-use change on macroinvertebrate communities of headwater streams in the Patagonian Andes. River Res. Appl. 2004, 20, 967–983. [Google Scholar] [CrossRef]
- Hepp, L.U.; Santos, S. Benthic communities of streams related to different land uses in a hydrographic basin in southern Brazil. Environ. Monit. Assess 2009, 157, 305–318. [Google Scholar] [CrossRef]
- Fu, L.; Jiang, Y.; Ding, J.; Liu, Q.; Peng, Q.-Z.; Kang, M.-Y. Impacts of land use and environmental factors on macroinvertebrate functional feeding groups in the Dongjiang River basin, southeast China. J. Freshw. Ecol. 2016, 31, 21–35. [Google Scholar] [CrossRef]
- Ley de Aguas Nacionales (L.A.N.). Comisión Nacional del Agua. Diario Oficial de la Federación. 01 de diciembre de 1992 México. Available online: http://www.diputados.gob.mx/LeyesBiblio/ref/lan.htm (accessed on 6 June 2018).



{kind=link}
{kind=link}
| Biological Metrics | Expected Response to Anthropogenic Activities |
|---|---|
| Category I “Taxonomic richness” | |
| Richness | Decrease |
| Rarified Richness | Decrease |
| Richness OCH (odonata + coleoptera + heteroptera) | Decrease |
| Richness EPT (ephemeroptera + plecoptera + trichoptera) | Decrease |
| Richness EPT/(richness EPT + richness OCH) | Decrease |
| Richness Ephemeroptera | Decrease |
| Richness Trichoptera | Decrease |
| Richness Odonata | Decrease |
| Richness Coleoptera | Decrease |
| Richness Diptera | Decrease |
| Richness Diptera wihout chironomidae | Decrease |
| Richness Gasteropoda | Decrease |
| Richness non-insect taxa (amphipoda + copepoda + ostracoda + gateropoda + oligochaeta + hirudinea) | Decrease |
| Category II “Taxonomic composition” | |
| % OCH (odonata + coleoptera + heteroptera) | Decrease |
| % EPT (ephemeroptera + plecoptera + trichoptera) | Decrease |
| % EPT/(% EPT + % OCH) | Decrease |
| % Ephemeroptera | Decrease |
| % Trichoptera | Decrease |
| % Odonata | Decrease |
| % Coleoptera | Decrease |
| % Diptera | Increase |
| % Diptera wihout chironomidae | Decrease |
| % Gasteropoda | Decrease |
| % Non-insect taxa (amphipoda + copepoda + ostracoda + gateropoda + oligochaeta + hirudinea) | Increase |
| % Baetidae/% EPT | Decrease |
| % Baetidae/% Ephemeroptera | Decrease |
| % Hydropsychidae/% EPT | Decrease |
| % Hydropsychidae/% Ephemeroptera | Decrease |
| % Chironomidae | Increase |
| % Oligochaeta | Increase |
| % Chironomidae + % Oligochaeta | Increase |
| Categoty III “Diversity indexes” | |
| Shannon’s Diversity using base e | Decrease |
| Simpson’s Diversity | Decrease |
| Pielou’s Evenness | Decrease |
| Category IV “Functional feeding groups” | |
| Richness Collectors-Gatherers | Decrease |
| % Collectors-Gatherers | Increase |
| Richness Predators | Decrease |
| % Predators | Decrease |
| Richness Herbivores | Decrease |
| % Herbivores | Decrease |
| Richness Collectors-Filterers | Decrease |
| % Collectors-Filterers | Decrease |
| ORDER | TAXA | CLASS | IV | p-Value |
|---|---|---|---|---|
| Odonata | Ophiogomphus | NAT | 1 | 0.001 |
| Trichoptera | Oxyethira | NAT | 1 | 0.001 |
| Hemiptera | Ambrysus | NAT | 0.99 | 0.001 |
| Ephemeroptera | Baetis | NAT | 0.90 | 0.001 |
| Diptera | Atrichpogon | NAT | 0.83 | 0.001 |
| Coleoptera | Macrelmis | NAT | 0.81 | 0.001 |
| Hemiptera | Rhagovelia | NAT | 0.78 | 0.001 |
| Trichoptera | Agryalea | NAT | 0.73 | 0.007 |
| Trichoptera | Chimarra | NAT | 0.67 | 0.002 |
| Coleoptera | Psephenus | NAT | 0.67 | 0.002 |
| Ephemeroptera | Farrodes | NAT | 0.66 | 0.004 |
| Ephemeroptera | Tricorythodes | NAT | 0.66 | 0.001 |
| Trichoptera | Mayatrichia | NAT | 0.66 | 0.002 |
| Megaloptera | Corydalus | NAT | 0.65 | 0.001 |
| Diptera | Limoniia | NAT | 0.65 | 0.002 |
| Diptera | Stratiomys | NAT | 0.65 | 0.001 |
| Diptera | Ceratopogon | NAT | 0.62 | 0.011 |
| Odonata | Nehalennia | NAT | 0.61 | 0.028 |
| Lepidoptera | Petrophila | NAT | 0.6 | 0.009 |
| Ephemeroptera | Caenis | NAT | 0.57 | 0.002 |
| Gasteropoda | M. tuberculata | NAT | 0.53 | 0.025 |
| Ephemeroptera | Camelobaetis | NAT | 0.50 | 0.003 |
| Coleoptera | Cymbiodita | NAT | 0.50 | 0.007 |
| Diptera | Hemerodromia | NAT | 0.50 | 0.008 |
| Odonata | Macrothemis | NAT | 0.46 | 0.007 |
| Coleoptera | Lutrochus | NAT | 0.46 | 0.013 |
| Ephemeroptera | Callibaetis | NAT | 0.45 | 0.007 |
| Trichoptera | Leucotrichia | NAT | 0.43 | 0.012 |
| Coleoptera | Paracymus | NAT | 0.33 | 0.035 |
| Diptera | A. fransiscanus | NAT | 0.33 | 0.039 |
| Diptera | Euparhypus | NAT | 0.33 | 0.023 |
| Coleoptera | Berosus | URB | 0.79 | 0.001 |
| Amphipoda | Hyallela azteca | AGR | 0.69 | 0.003 |
| Hirudinea | Erpobdellidae | AGR | 0.64 | 0.002 |
| Diptera | Chironomus | AGR | 0.45 | 0.012 |
| Odonata | Ischnura | AGR | 0.36 | 0.029 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-López, D.; Rodríguez-Lozano, P.; Arias-Real, R.; Guerra-Cobián, V.; Prat, N. The Influence of Riparian Corridor Land Use on the Pesquería River’s Macroinvertebrate Community (N.E. Mexico). Water 2019, 11, 1930. https://doi.org/10.3390/w11091930
Castro-López D, Rodríguez-Lozano P, Arias-Real R, Guerra-Cobián V, Prat N. The Influence of Riparian Corridor Land Use on the Pesquería River’s Macroinvertebrate Community (N.E. Mexico). Water. 2019; 11(9):1930. https://doi.org/10.3390/w11091930
Chicago/Turabian StyleCastro-López, Daniel, Pablo Rodríguez-Lozano, Rebeca Arias-Real, Víctor Guerra-Cobián, and Narcís Prat. 2019. "The Influence of Riparian Corridor Land Use on the Pesquería River’s Macroinvertebrate Community (N.E. Mexico)" Water 11, no. 9: 1930. https://doi.org/10.3390/w11091930