
Cyclic Peptide-Gadolinium Nanocomplexes as siRNA Delivery Tools

 , and
, and 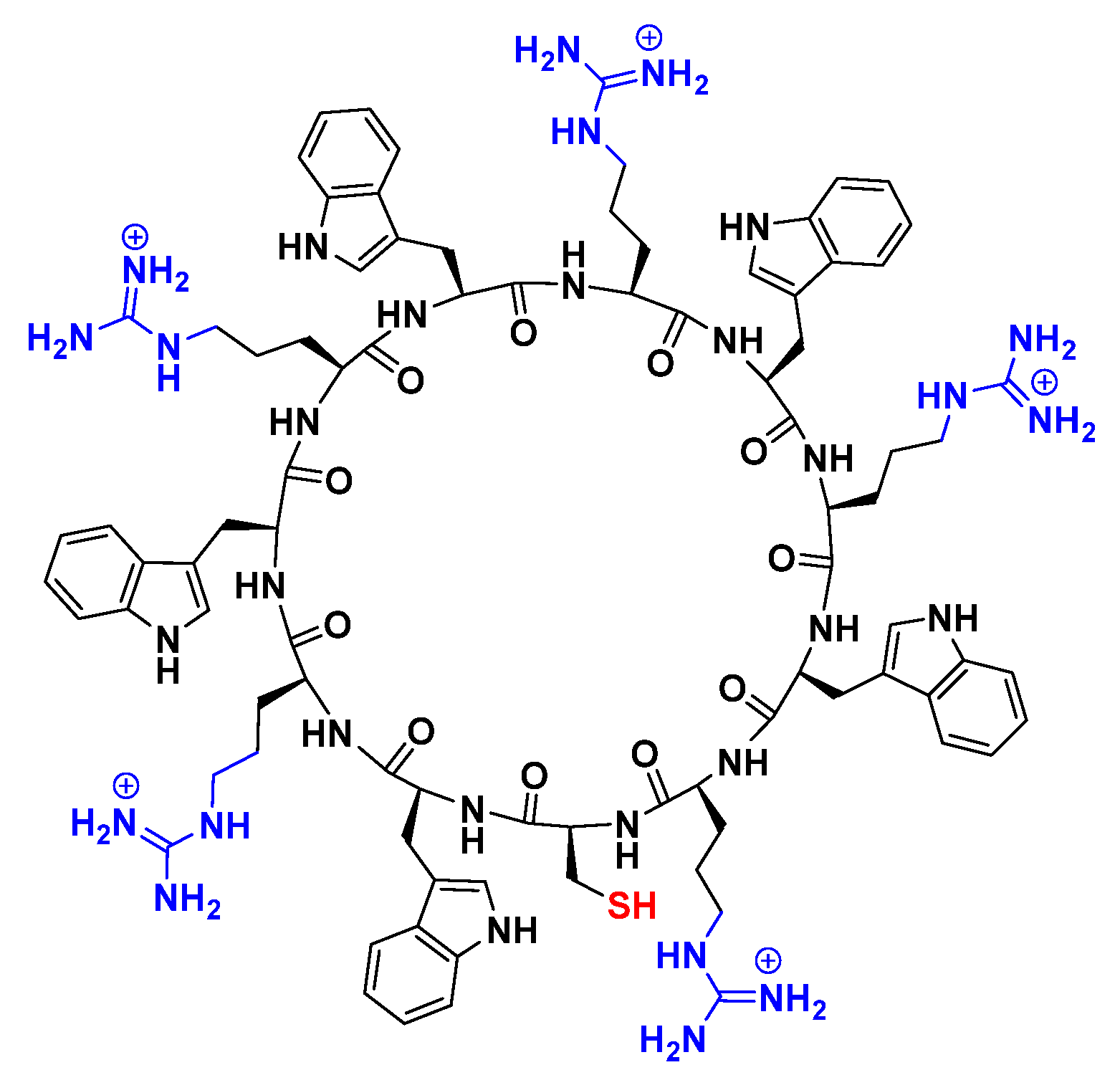{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussions
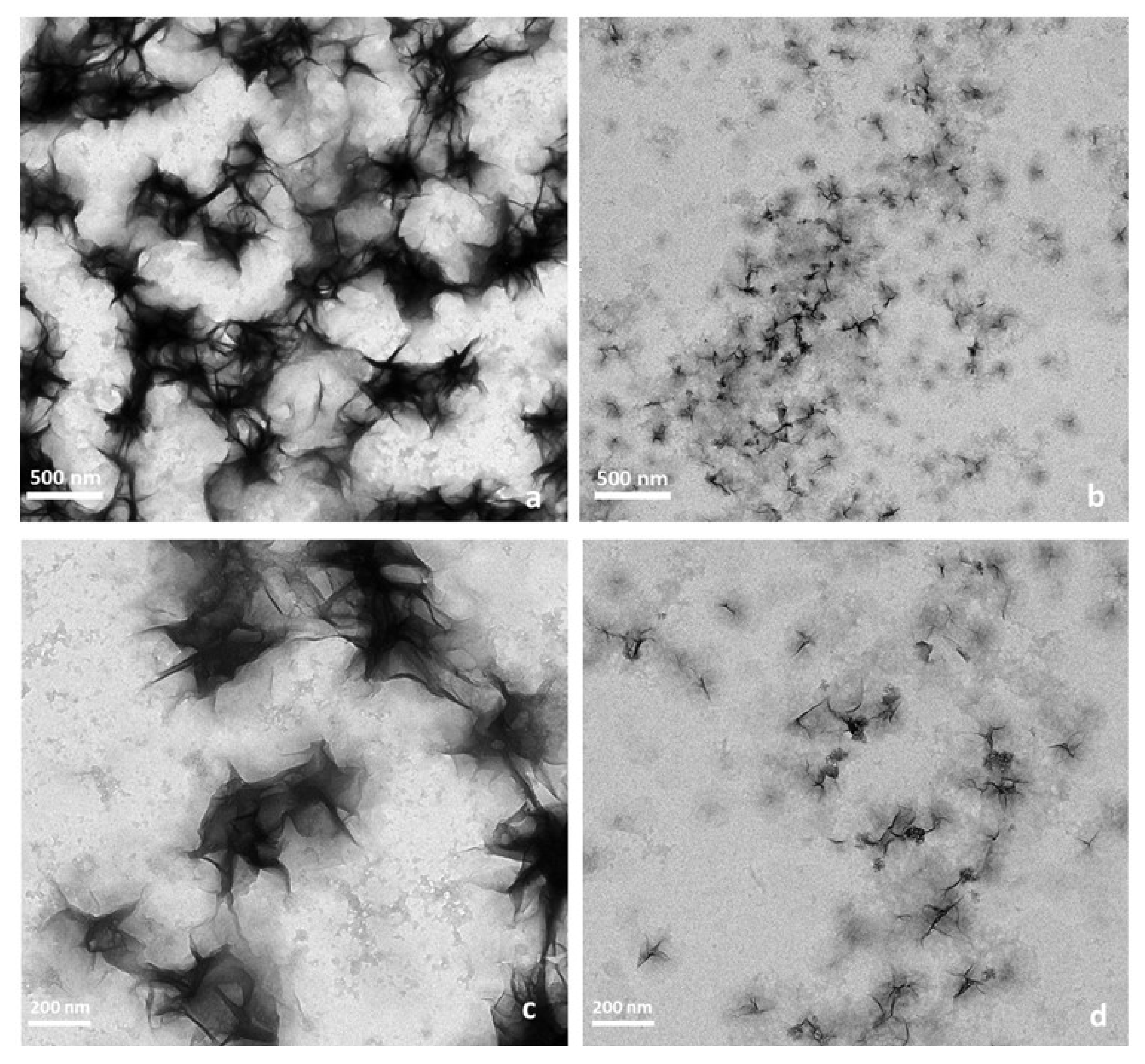
2.1. Preparation and Characterization of Peptide-GdNPs
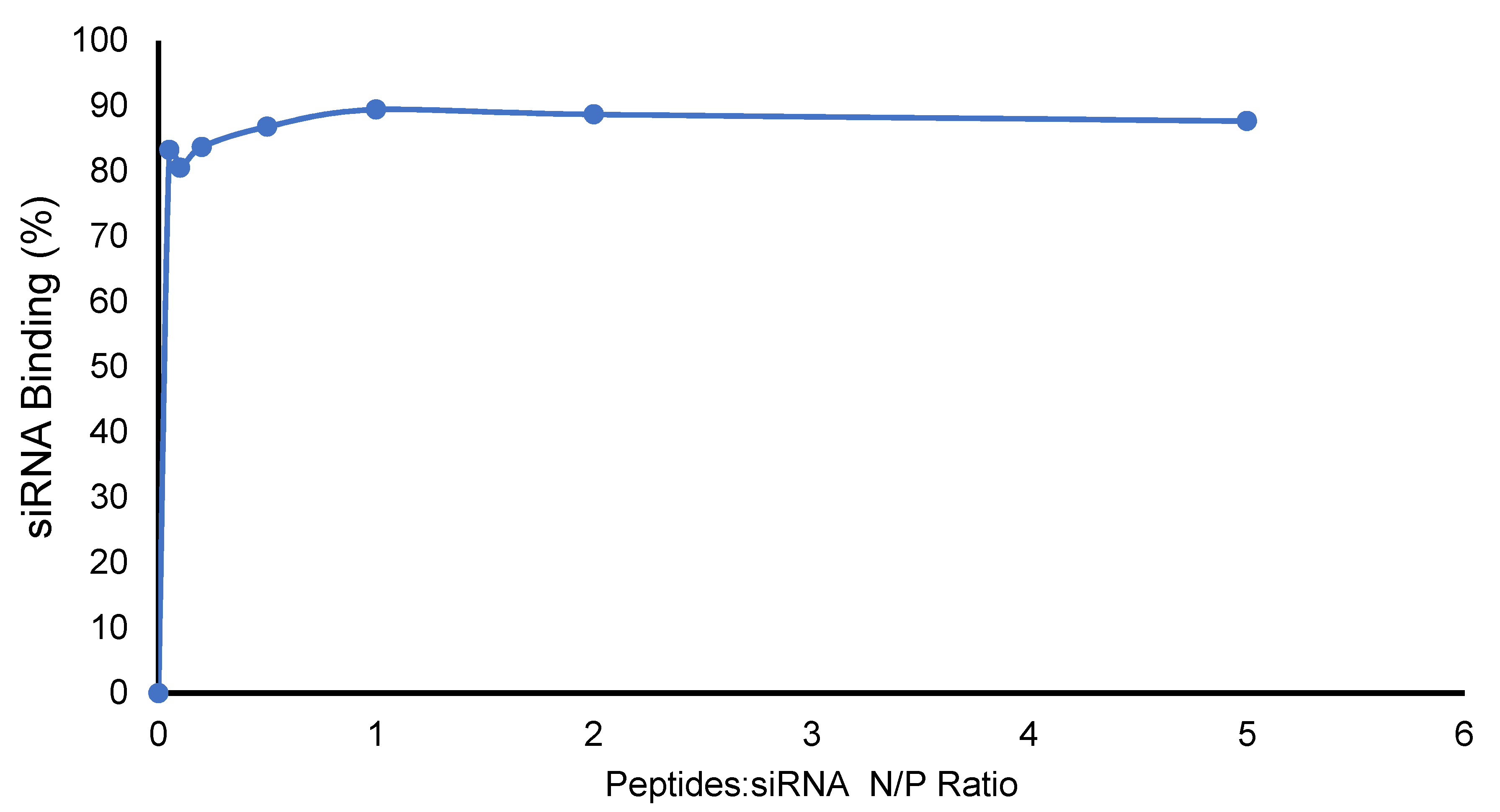
2.2. Formation of Peptide-–Nanoparticles and siRNA Complexes and Their Binding Affinity
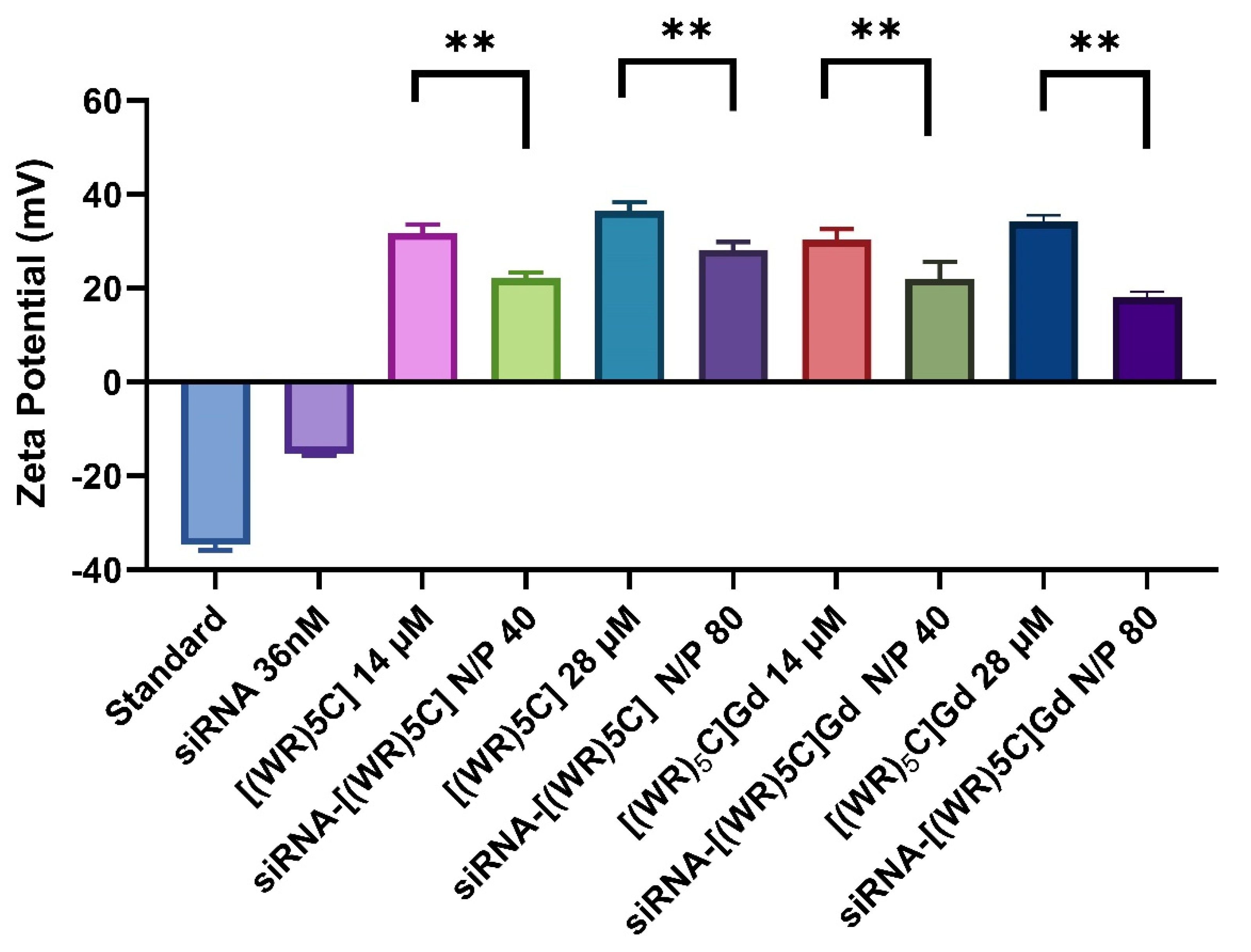
2.3. Zeta Potential of Peptide-GdNPs
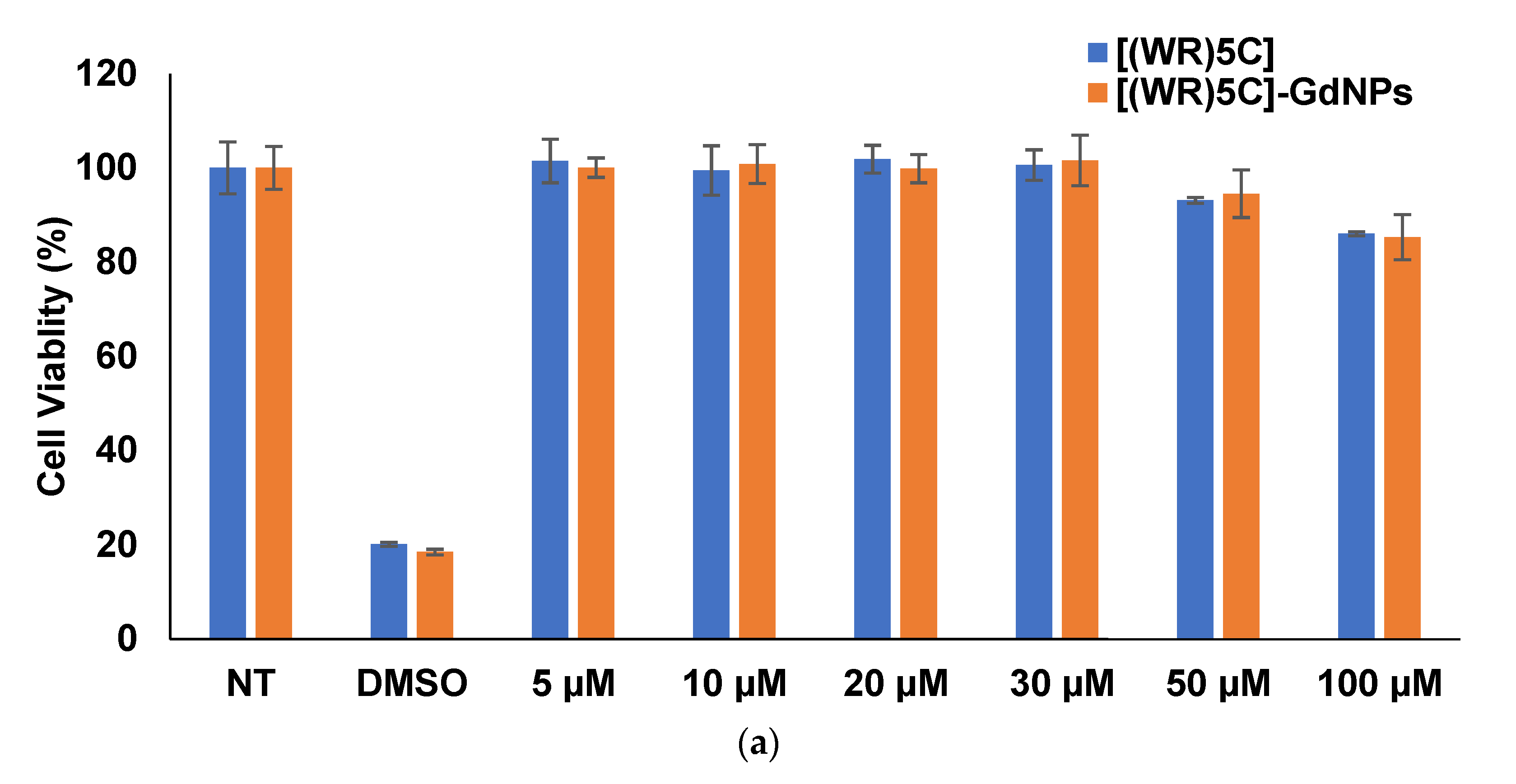
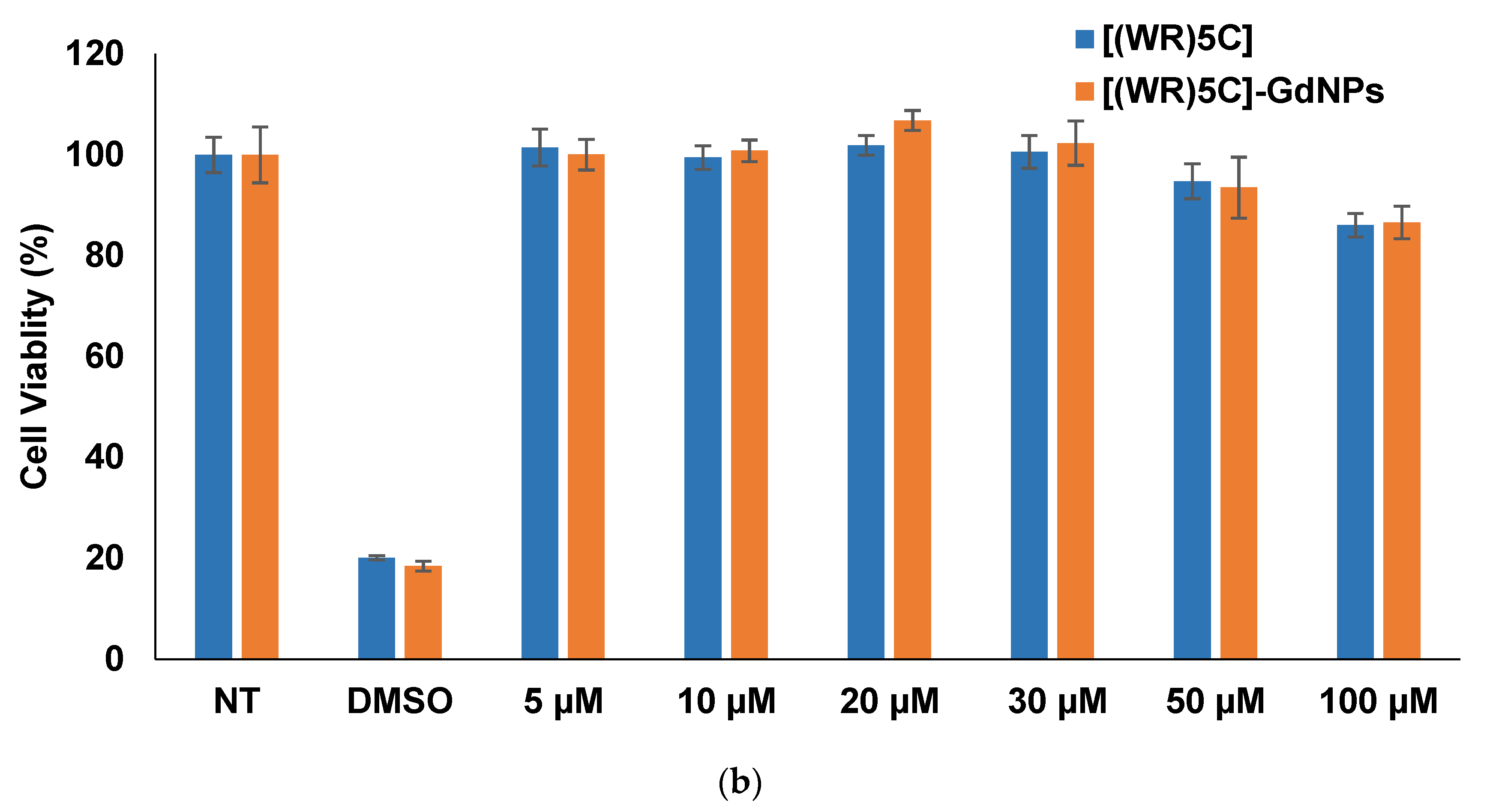
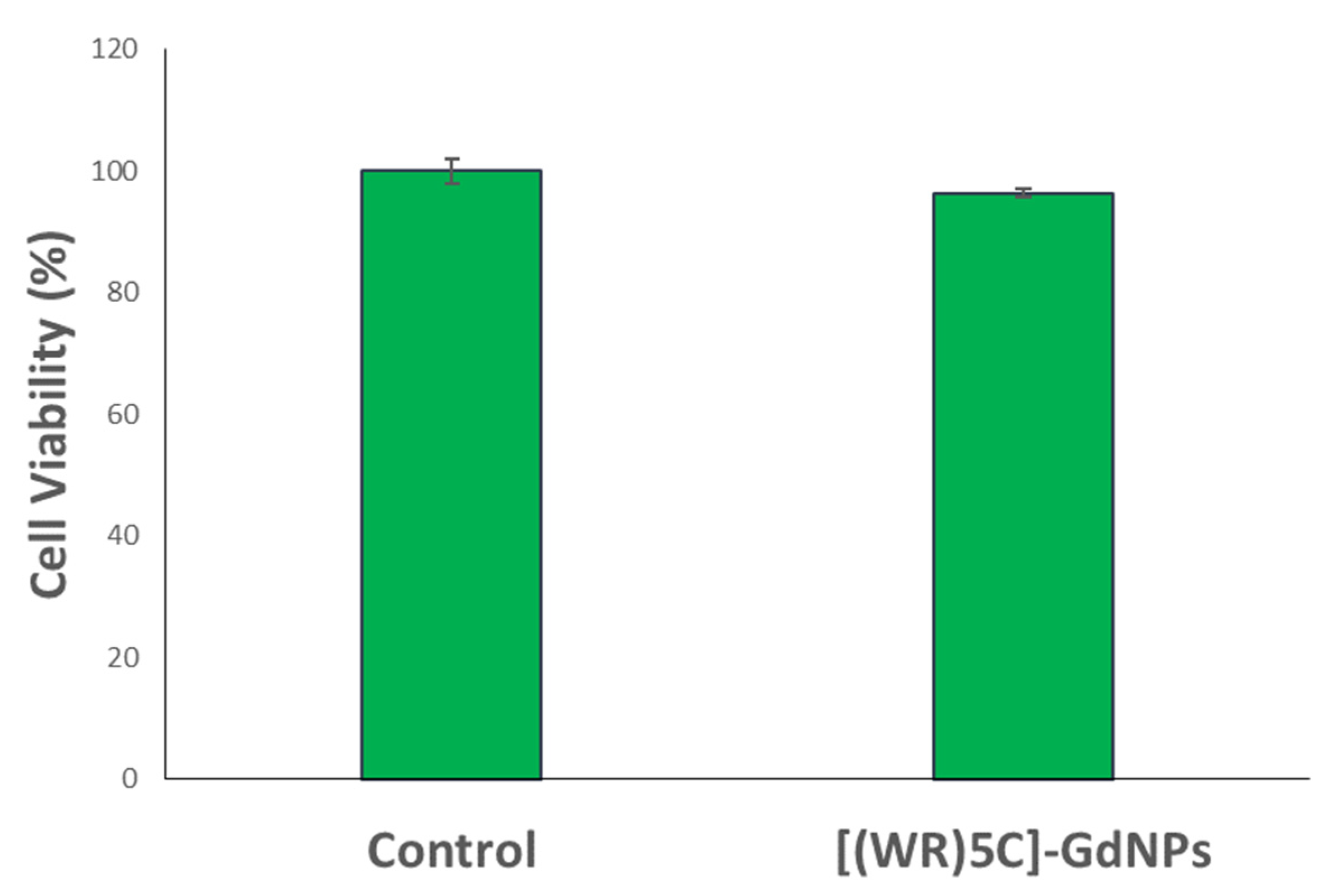
2.4. Cytotoxicity of Peptide-GdNPs
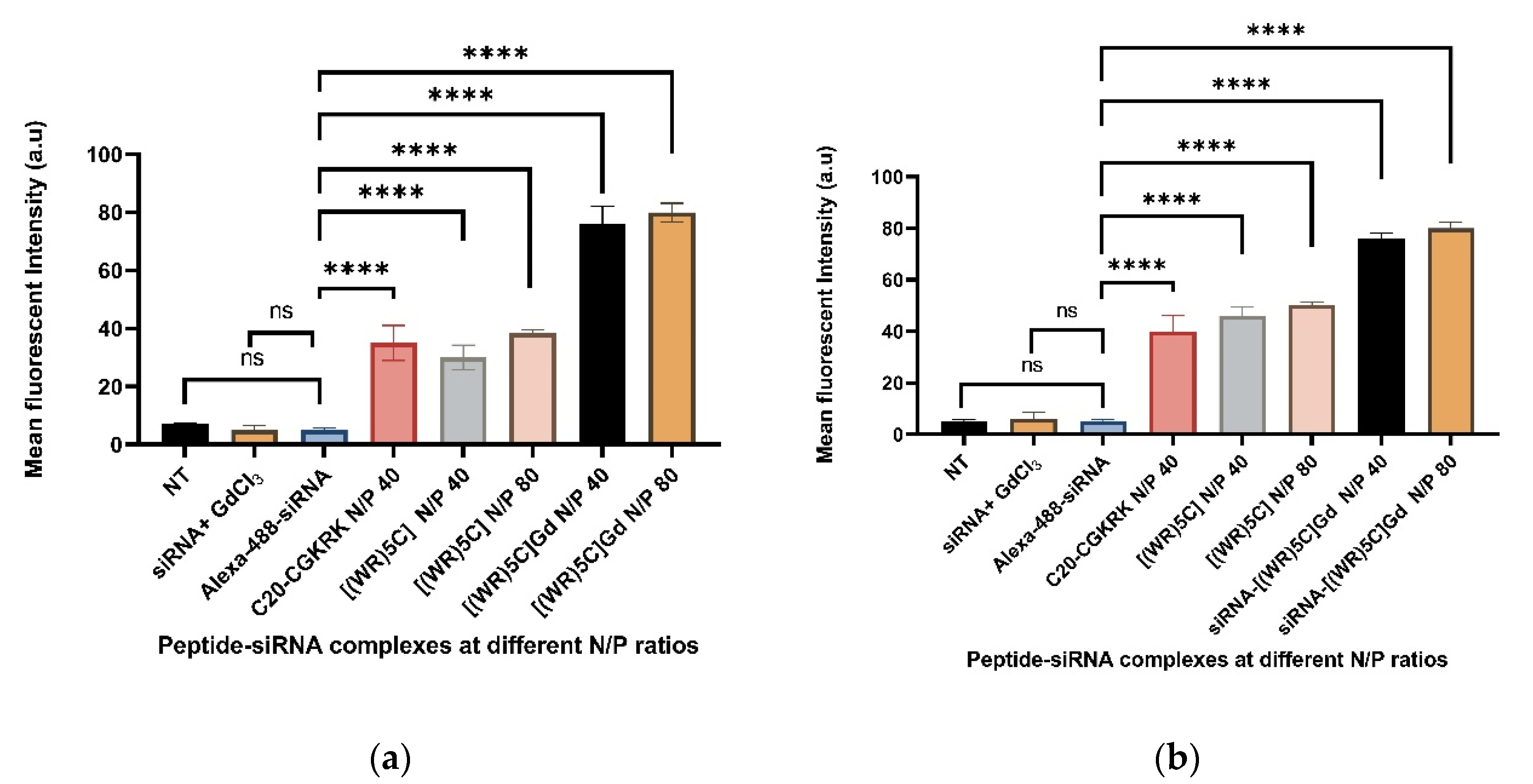
2.5. Evaluation of Peptide-GdNPs as Molecular Transporters
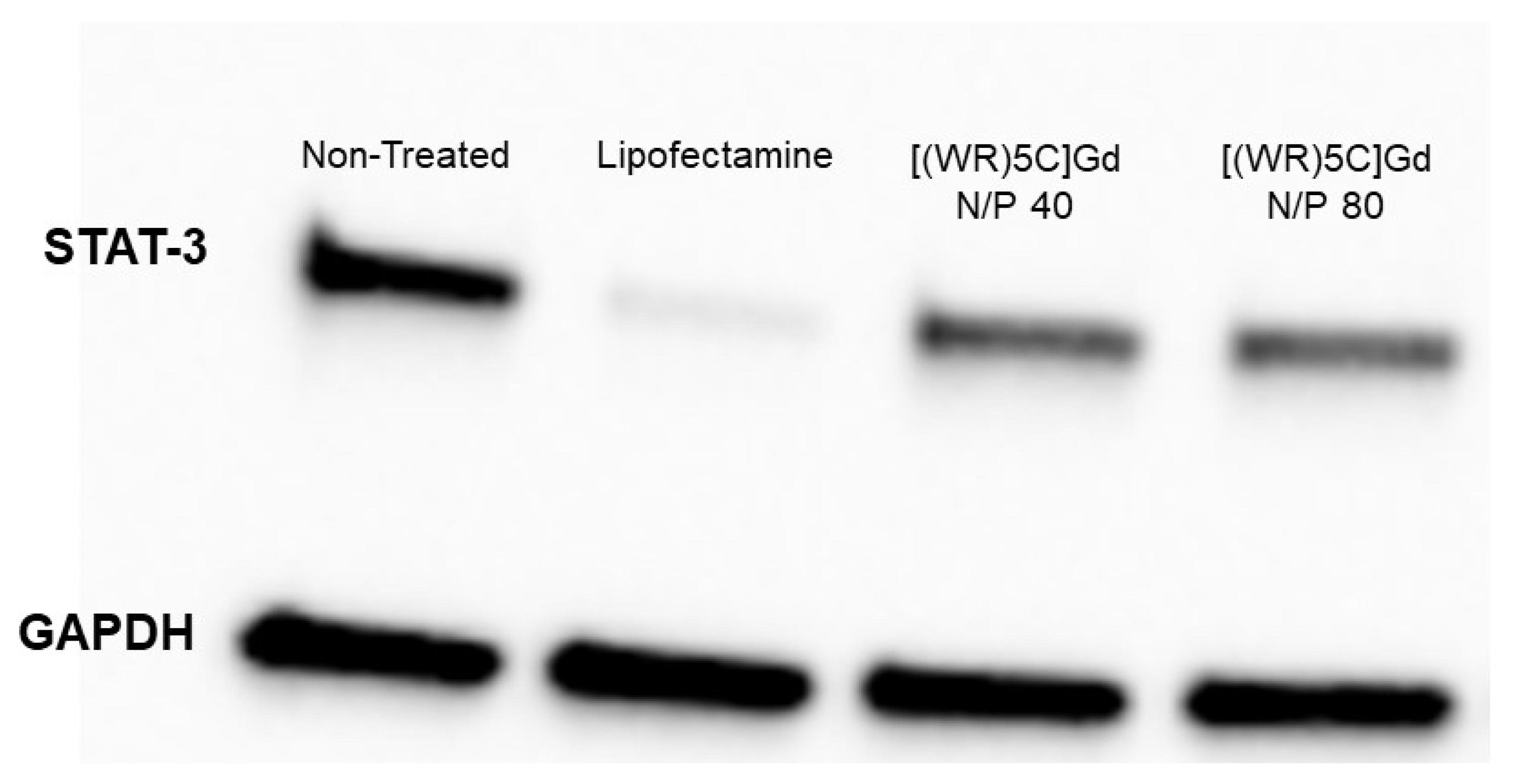
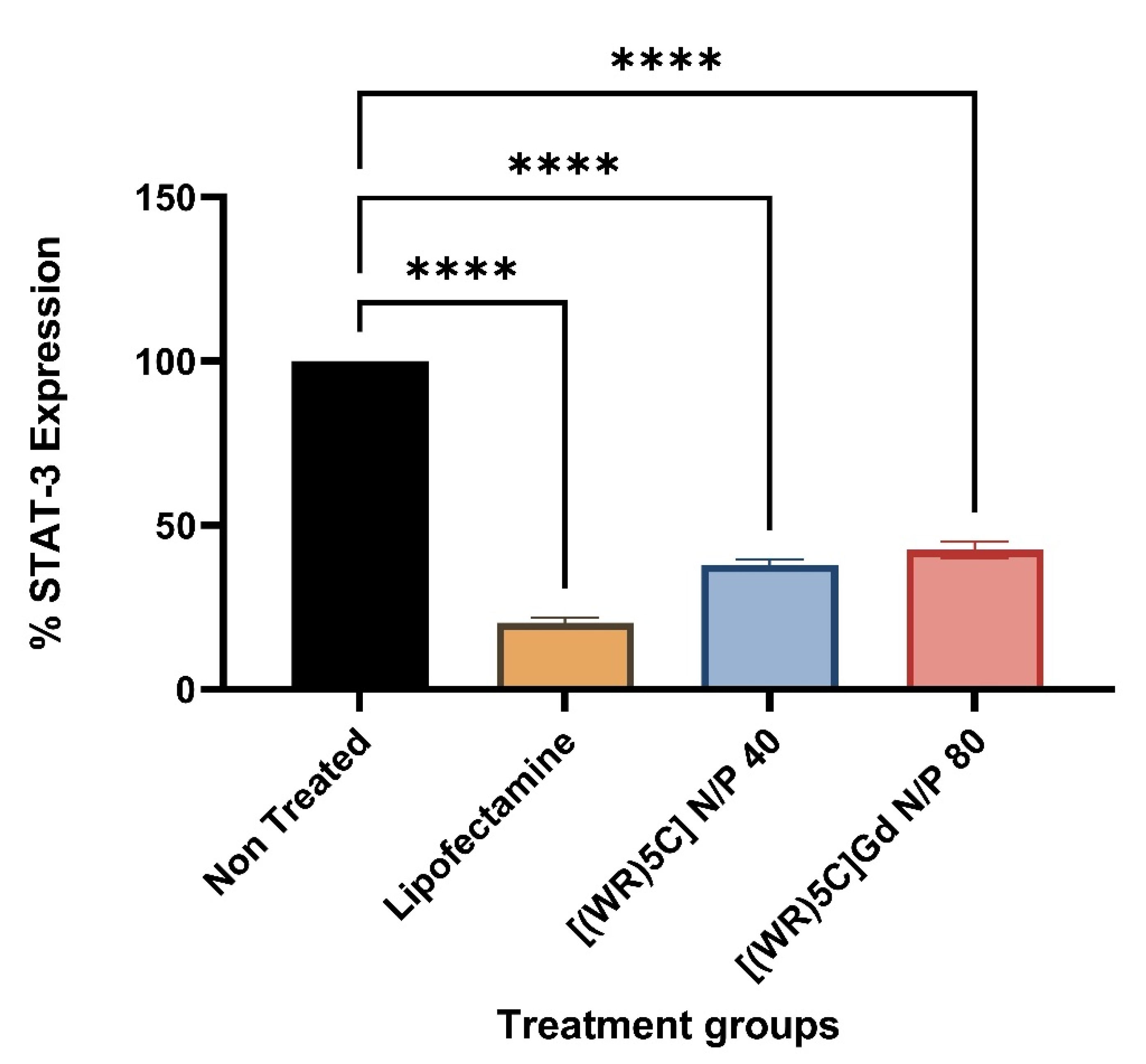
2.6. Protein Quantification (Western Blot)
2.7. Membrane Integrity
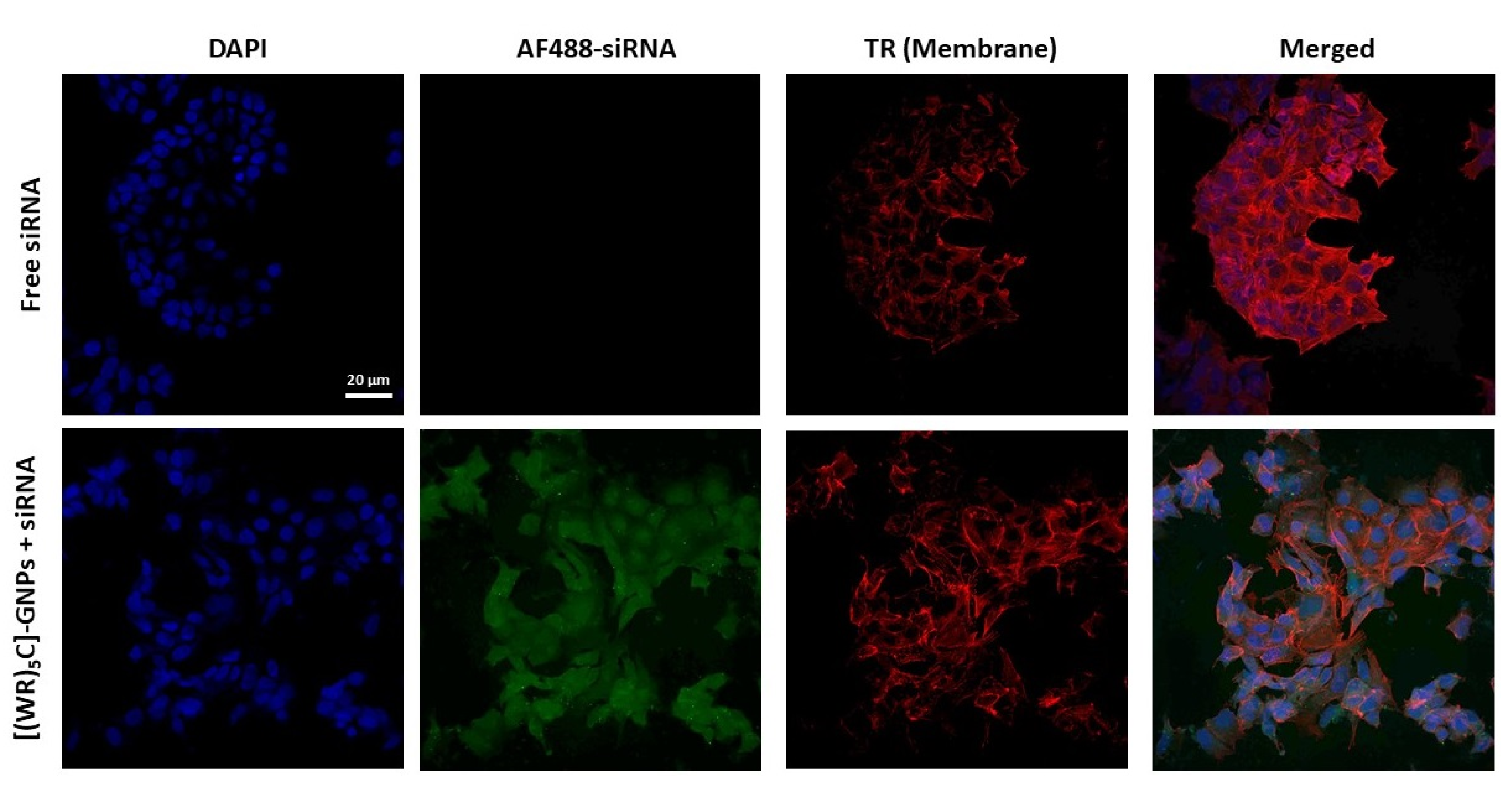
2.8. Confocal Microscopy Imaging
3. Materials and Methods
3.1. Synthesis of Cyclic Peptide [CWRWRWRWRWR] (1)
3.2. In Situ Preparation of Cyclic [(WR)5C]-GdNPs
3.3. Complex Formation of Cyclic [(WR)5C]-GdNPs/siRNA Complexes, Binding Affinity, and BC50
3.4. Zeta Potential
3.5. Transmission Electron Microscopy (TEM)
3.6. Cell Culture
3.7. Cytotoxicity Assay of [(WR)5C]-GdNPs
3.8. Flow Cytometry Studies
3.9. Protein Quantification (Western Blot)
3.10. Membrane Integrity Test
3.11. Confocal Microscopy Imaging
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Sontheimer, E.J. Assembly and function of RNA silencing complexes. Nat. Rev. Mol. Cell Biol. 2005, 6, 127–138. [Google Scholar] [CrossRef]
- Sahay, G.; Querbes, W.; Alabi, C.; Eltoukhy, A.; Sarkar, S.; Zurenko, C.; Karagiannis, E.; Love, K.; Chen, D.; Zoncu, R.; et al. Efficiency of siRNA delivery by lipid nanoparticles is limited by endocytic recycling. Nat. Biotechnol. 2013, 31, 653–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilleron, J.; Querbes, W.; Zeigerer, A.; Borodovsky, A.; Marsico, G.; Schubert, U.; Manygoats, K.; Seifert, S.; Andree, C.; Stöter, M.; et al. Image-based analysis of lipid nanoparticle-mediated siRNA delivery, intracellular trafficking and endosomal escape. Nat. Biotechnol. 2013, 31, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Lehto, T.; Ezzat, K.; Wood, M.J.; EL Andaloussi, S. Peptides for nucleic acid delivery. Adv. Drug Deliv. Rev. 2016, 106, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Tai, W.; Gao, X. Functional peptides for siRNA delivery. Adv. Drug Deliv. Rev. 2016, 110, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Järver, P.; Mäger, I.; Langel, Ü. In vivo biodistribution and efficacy of peptide mediated delivery. Trends Pharmacol. Sci. 2010, 31, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Layek, B.; Lipp, L.; Singh, J. Cell Penetrating Peptide Conjugated Chitosan for Enhanced Delivery of Nucleic Acid. Int. J. Mol. Sci. 2015, 16, 28912–28930. [Google Scholar] [CrossRef] [Green Version]
- Bellich, B.; D’Agostino, I.; Semeraro, S.; Gamini, A.; Cesàro, A. The good, the bad and the ugly of chitosans. Mar. Drugs 2016, 14, 99. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.P.; Patel, R.R.; Patel, J.K. Chitosan Mediated Targeted Drug Delivery System: A Review. J. Pharm. Pharm. Sci. 2010, 13, 536–557. [Google Scholar] [CrossRef]
- Singha, K.; Namgung, R.; Kim, W. Polymers in small-interfering RNA delivery. Nucleic Acid Ther. 2011, 21, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Torchilin, V.P. Dendrimers for siRNA Delivery. Pharmaceuticals 2013, 6, 161–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannan, R.M.; Nance, E.; Kannan, S.; Tomalia, D.A. Emerging concepts in dendrimer-based nanomedicine: From design principles to clinical applications. J. Intern. Med. 2014, 276, 579–617. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Gemeinhart, R.A. Progress in microRNA delivery. J. Control. Release 2013, 172, 962–974. [Google Scholar] [CrossRef] [Green Version]
- Kozielski, K.L.; Tzeng, S.Y.; Green, J.J. Bioengineered nanoparticles for siRNA delivery. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnology 2013, 5, 449–468. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Jiang, Z.; Saha, K.; Kim, C.S.; Kim, S.T.; Landis, R.F.; Rotello, V.M. Gold Nanoparticles for Nucleic Acid Delivery. Mol. Ther. 2014, 22, 1075–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, S.; Bajaj, A.; Mout, R.; Rotello, V.M. Monolayer coated gold nanoparticles for delivery applications. Adv. Drug Deliv. Rev. 2012, 64, 200–216. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Duan, J.; Wang, J.; Liu, Q.; Shang, R.; Yang, X.; Lu, P.; Xia, C.; Wang, L.; Dou, K. Superparamagnetic iron oxide nanoparticles modified with polyethylenimine and galactose for siRNA targeted delivery in hepatocellular carcinoma therapy. Int. J. Nanomed. 2018, 13, 1851–1865. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, N.S.; Tekade, R.K.; Chougule, M.B. Nanocarrier mediated delivery of siRNA/miRNA in combination with chemotherapeutic agents for cancer therapy: Current progress and advances. J. Control. Release 2014, 194, 238–256. [Google Scholar] [CrossRef] [Green Version]
- Lin, G.; Chen, T.; Zou, J.; Wang, Y.; Wang, X.; Li, J.; Huang, Q.; Fu, Z.; Zhao, Y.; Lin, M.-C.; et al. Quantum dots-siRNA nanoplexes forgene silencing in central nervoussystem tumor cells. Front. Pharmacol. 2017, 8, 182. [Google Scholar] [CrossRef] [Green Version]
- Singh, T.; Murthy, A.S.N.; Yang, H.-J.; Im, J. Versatility of cell-penetrating peptides for intracellular delivery of siRNA. Drug Deliv. 2018, 25, 1996–2006. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Ding, F.; Zhang, J.; Guo, Y.; Gao, X.; Feng, J.; Zhu, X.; Zhang, C. DNA tetrahedron-based nanogels for siRNA delivery and gene silencing. Chem. Commun. 2019, 55, 4222–4225. [Google Scholar] [CrossRef]
- Garber, K. Alnylam launches era of RNAi drugs. Nat. Biotechnol. 2018, 36, 777–778. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food Drug Administration. Information on Gadolinium-Based Contrast Agents. Available online: https://www.fda.gov/drugs/postmarket-drug-safety-information-patients-and-providers/information-gadolinium-based-contrast-agents (accessed on 20 August 2020).
- Cho, H.K.; Cho, H.-J.; Lone, S.; Kim, D.-D.; Yeum, J.H.; Cheong, I.W. Preparation and characterization of MRI-active gadolinium nanocomposite particles for neutron capture therapy. J. Mater. Chem. 2011, 21, 15486–15493. [Google Scholar] [CrossRef]
- Gulani, V.; Calamante, F.; Shellock, F.G.; Kanal, E.; Reeder, S.B. Gadolinium deposition in the brain: Summary of evidence and recommendations. Lancet Neurol. 2017, 16, 564–570. [Google Scholar] [CrossRef]
- Na, K.; Lee, S.A.; Jung, S.H.; Shin, B.C. Gadolinium-based cancer therapeutic liposomes for chemotherapeutics and diagnostics. Colloids Surf. B Biointerfaces 2011, 84, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Frullano, L.; Tejerina, B.; Meade, T.J. Synthesis and characterization of a doxorubicin−Gd(III) contrast agent conjugate: A new approach toward prodrug−procontrast complexes. Inorg. Chem. 2006, 45, 8489–8491. [Google Scholar] [CrossRef]
- Wang, L.; Xing, H.; Zhang, S.; Ren, Q.; Pan, L.; Zhang, K.; Bu, W.; Zheng, X.; Zhou, L.; Peng, W.; et al. A Gd-doped Mg-Al-LDH/Au nanocomposite for CT/MR bimodal imagings and simultaneous drug delivery. Biomaterials 2013, 34, 3390–3401. [Google Scholar] [CrossRef]
- Park, S.E.; Sajid, M.I.; Parang, K.; Tiwari, R.K. cyclic cell-penetrating peptides as efficient intracellular drug delivery tools. Mol. Pharm. 2019, 16, 3727–3743. [Google Scholar] [CrossRef]
- Do, H.; Sharma, M.; El-Sayed, N.S.; Mahdipoor, P.; Bousoik, E.; Parang, K.; Aliabadi, H.M. Difatty acyl-conjugated linear and cyclic peptides for siRNA delivery. ACS Omega 2017, 2, 6939–6957. [Google Scholar] [CrossRef]
- Sharma, M.; El-Sayed, N.S.; Do, H.; Parang, K.; Tiwari, R.K.; Aliabadi, H.M. Tumor-targeted delivery of siRNA using fatty acyl-CGKRK peptide conjugates. Sci. Rep. 2017, 7, 6093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirazi, A.N.; Mozaffari, S.; Sherpa, R.T.; Tiwari, R.; Parang, K. Efficient intracellular delivery of cell-impermeable cargo molecules by peptides containing tryptophan and histidine. Molecules 2018, 23, 1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirazi, A.N.; Tiwari, R.; Chhikara, B.S.; Mandal, D.; Parang, K. Design and biological evaluation of cell-penetrating peptide–doxorubicin conjugates as prodrugs. Mol. Pharm. 2013, 10, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Mandal, D.; Shirazi, A.N.; Parang, K. Cell-Penetrating Homochiral Cyclic Peptides as Nuclear-Targeting Molecular Transporters. Angew. Chem. Int. Ed. 2011, 50, 9633–9637. [Google Scholar] [CrossRef] [PubMed]
- Shirazi, A.N.; Paquin, K.L.; Howlett, N.G.; Mandal, D.; Parang, K. Cyclic peptide-capped gold nanoparticles for enhanced siRNA delivery. Molecules 2014, 19, 13319–13331. [Google Scholar] [CrossRef]
- Shirazi, A.N.; Tiwari, R.K.; Oh, D.; Sullivan, B.; McCaffrey, K.; Mandal, D.; Parang, K. Surface decorated gold nanoparticles by linear and cyclic peptides as molecular transporters. Mol. Pharm. 2013, 10, 3137–3151. [Google Scholar] [CrossRef] [Green Version]
- Shirazi, A.N.; Tiwari, R.K.; Oh, D.; Sullivan, B.; Kumar, A.; Beni, Y.A.; Parang, K. Cyclic Peptide–Selenium Nanoparticles as Drug Transporters. Mol. Pharm. 2014, 11, 3631–3641. [Google Scholar] [CrossRef]
- Shirazi, A.N.; Park, S.; Rad, S.; Baloyan, L.; Mandal, D.; Sajid, M.; Hall, R.; Lohan, S.; Zoghebi, K.; Parang, K.; et al. Cyclic peptide-gadolinium nanoparticles for enhanced intracellular delivery. Pharmaceutics 2020, 12, 792. [Google Scholar] [CrossRef]
- Mozaffari, S.; Bousoik, E.; Amirrad, F.; Lamboy, R.; Coyle, M.; Hall, R.; Alasmari, A.; Mahdipoor, P.; Parang, K.; Aliabadi, H.M. Amphiphilic peptides for efficient siRNA delivery. Polymers 2019, 11, 703. [Google Scholar] [CrossRef] [Green Version]
- Panigrahi, B.; Singh, R.K.; Mishra, S.; Mandal, D. Cyclic peptide-based nanostructures as efficient siRNA carriers. Artif. Cells Nanomed. Biotechnol. 2018, 46, S763–S773. [Google Scholar] [CrossRef] [Green Version]
- Grayson, A.C.R.; Doody, A.M.; Putnam, D. Biophysical and structural characterization of polyethylenimine-mediated siRNA delivery in vitro. Pharm. Res. 2006, 23, 1868–1876. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nasrolahi Shirazi, A.; Sajid, M.I.; Mandal, D.; Stickley, D.; Nagasawa, S.; Long, J.; Lohan, S.; Parang, K.; Tiwari, R.K. Cyclic Peptide-Gadolinium Nanocomplexes as siRNA Delivery Tools. Pharmaceuticals 2021, 14, 1064. https://doi.org/10.3390/ph14111064
Nasrolahi Shirazi A, Sajid MI, Mandal D, Stickley D, Nagasawa S, Long J, Lohan S, Parang K, Tiwari RK. Cyclic Peptide-Gadolinium Nanocomplexes as siRNA Delivery Tools. Pharmaceuticals. 2021; 14(11):1064. https://doi.org/10.3390/ph14111064
Chicago/Turabian StyleNasrolahi Shirazi, Amir, Muhammad Imran Sajid, Dindyal Mandal, David Stickley, Stephanie Nagasawa, Joshua Long, Sandeep Lohan, Keykavous Parang, and Rakesh Kumar Tiwari. 2021. "Cyclic Peptide-Gadolinium Nanocomplexes as siRNA Delivery Tools" Pharmaceuticals 14, no. 11: 1064. https://doi.org/10.3390/ph14111064